|
УДК 630*561.24: 630*453
Павлов И.Н., Агеев А.А.
Влияние зоогенной дефолиации на структуру годичного слоя древесины сосны обыкновенной
Установлены закономерности ответной реакции ксилемы Pinus sylvestrisна дефолиацию DendrolimussuperanssibiricusTschetv. Восстановление хвои сопровождается адекватным ростом ширины годичного слоя. При этом увеличению радиального прироста предшествует рост площади трахеид. Данная закономерность характерна только для периода интенсивной дефолиации и может быть использована для идентификации вспышек массового размножения филофагов.
Панзональные вспышки массового размножения сибирского шелкопряда (СШ) являются фактором, во многом определяющим рост и распространение хвойных лесов Северной Азии. В подзоне южной тайги Западной Сибири (бассейны рек Чулыма, Кемчуга, Кети и Кеми) в результате деятельности сибирского шелкопряда в 1952 - 1957 г.г. усохло около 1,5 млн. га темнохвойных лесов (Фуряев, 1966). Его особая вредоносность заключается в полном уничтожении в очагах темнохвойного древостоя (или лиственницы), включая возобновление, а также следующих за этим пожаров (Дудин 1958, Фалалеев, 1960; Коломиец, 1962; Крылов и др., 1975; Исаев и др., 2001). Из-за этого в погибших древостоях в большинстве случаев отсутствует удовлетворительное возобновление не только хвойных, но и лиственных пород.
Для сибирского шелкопряда характерна определенная периодичность вспышек массового размножения. Достаточно часто дефолиации в разной степени подвергаются одни и те же насаждения. Изучение закономерностей возникновения вспышек, их цикличности, а также развития лесной растительности после массового размножения сибирского шелкопряда представляет значительный научный и практический интерес. Одним из способов получения информации о прошлых событиях является дендрохронология. Интенсивная дефолиация сопровождается значительной редукцией радиального прироста (Ваганов, Терсков, 1977; Федоренко, 1997; Ваганов, Шашкин, 2000; Свидерская, Пальникова, 2003; Seija et al., 2002; Hogg et al., 2002).
Однако снижение ширины годичного кольца может произойти также в случае неблагоприятного сочетания погодных факторов, пожаров, изменения ценотических условий. Кроме всего прочего, в связи с неравномерным ростом по периметру ствола объективную информацию о текущем приросте можно получить только по нескольким кернам с одного дерева (не менее 4). Следовательно, необходимо найти маркерные признаки, отличающиеся малой изменчивостью в период действия изучаемого фактора. Тогда, где бы мы не взяли керн - всегда сможем идентифицировать время вспышки массового размножения СШ.
Объедание хвои насекомыми, как и подрезка ветвей, в первую очередь отражается на радиальном приросте ствола и лишь после более сильного объедания - на линейном приросте побегов (Лир и др., 1974). Прирост побегов поддерживается за счет снижения прироста древесины и достигает минимума на 1-3 года позднее радиального прироста ствола (Рожков, Хлиманкова, Степанчук , 1991).
В год дефолиации лиственницы (искусственной или вызванной объеданием хвои насекомыми) уменьшается ширина годичного кольца, в поздней зоне формируются клетки с тонкими стенками, уменьшаются размеры клеточного люмена, уменьшается плотность годичных колец (Ваганов, Терсков, 1977; Ваганов, Шашкин, 2000; Schweingruber, 1979; Filion, Cournoyer, 1995).
А.С. Рожков и др. (1991) отмечают, что, несмотря на очевидную тенденцию, даже при критическом состоянии дерева, к подержанию свойственного породе соотношения ранней и поздней древесины (ксилемный гомеостаз), у ели и кедра после сильной дефолиации количество рядов толстостенных трахеид увеличивается в 2-3 раза. Более сложный характер ответной реакции лиственницы на потерю хвои в результате атаки гусениц шелкопряда отмечают Е.А. Ваганов и И.А. Терсков (1977). Первоначально, помимо уменьшения ширины годичных колец и доли поздней древесины, изменяются показатели более детального строения ксилемы; несколько уменьшается диаметр ранних трахеид на всех высотах ствола и происходит редукция толщины их стенок. Более заметные отклонения от нормы наблюдаются в зоне трахеид поздней древесины: резко сокращается толщина стенок клеток. На следующий год значительно изменяется соотношение между количеством ранней и поздней древесины в пользу последней (Ваганов, Терсков, 1977). Е.С. Чавчавадзе (1979) и Н.Е. Косиченко (1999) связываютуменьшение ширины годичного кольца у хвойных, вызванное угнетением, прежде всего с сокращением объема ранней древесины В целом, у хронически ослабленных деревьев при предельном сокращении прироста годичное кольцо может состоять из одного ряда тонкостенных и одного толстостенных трахеид или только из одного ряда толстостенных (Рожков, Хлиманкова , Степанчук , 1991).
При хроническом атмосферном загрязнении надежными показателями состояния ели сибирской и сосны обыкновенной являются: толщина тангентальных стенок трахеид ранней и поздней древесины; радиальный диаметр ранних трахеид; ширина годичного кольца (Арсеньева, Чавчавадзе, 2001). При увеличении техногенной нагрузки наблюдается устойчивое снижение указанных биометрических показателей. Радиальный диаметр просветов трахеид ранней древесины ели сибирской не поддается какой-либо закономерности - он почти не меняется с ухудшением условий произрастания (Арсеньева, Чавчавадзе, 2001).
В целом, влияние дефолиации на структуру годичных колец можно классифицировать как опосредованное, оказывающее действие через уровень ассимилятов и гормонов роста (Ваганов, Шашкин, 2000).
Объекты и методы исследования
Закономерности возобновления и формирования леса изучали на территории Средне-Чулымского лесничества Тегульдетского лесхоза. Согласно лесорастительному районированию Западной Сибири, территория исследования относится к южной части зоны средней тайги, к Чулымо-Енисейскому сосново-темнохвойному округу (Крылов, 1960). Особая уникальность района исследования состоит в близком соседстве почв разного гранулометрического состава (песок, супесь, суглинок, глина).
Исследуемая территория была подвержена панзональным вспышкам размножения сибирского шелкопряда в 1914-1922 (Дудин, 1958; Коломиец, 1962); 1920 -1928 (Коломиец, Майер, 1963); 1950—1959 гг. (Катаев, 1959; Коломиец, 1962).
Вспышка 1950—1959 гг. сформировалась в центральной части Красноярского края и в Томской области в условиях заболоченной темнохвойной тайги, где до этого развитие очагов хвоегрызущих насекомых считалось маловероятным (Коломиец, 1962). В 1954 г. очаги шелкопряда были обнаружены в Средне-Чулымском лесничестве Тегульдетского лесхоза, где вспышка началась в древостоях, произрастающих вдоль реки Чичка-Юл (Катаев, 1959).
Во время экспедиции 2002 г. для исследования закономерностей радиального прироста по общепринятым методикам были взяты керны с единичных деревьев сосны обыкновенной, переживших вспышку массового размножения СШ. Для гистометрического анализа были выбраны 9 деревьев одного класса возраста (для устранения влияния возраста меристемы на продуцирование трахеид) с ярко выраженной редукцией прироста. Для выявления характерных при зоогенной дефолиации закономерностей образования ксилемы в качестве контроля для исследования было взято 9 деревьев из района гарантированного отсутствия объедания хвои СШ.
Фотографии поперечных срезов были сделаны на микроскопах: электронном сканирующем РЭМ 100-У и Оlympus СХ-41 с фотокамерой Nicon coolpix 4500 при х400 увеличении (рис.1). На полученных изображениях с помощью разработанной компьютерной программы (Павлов и др., 2005) произведены измерения основных биометрических показателей трахеид (площади клеток и их люменов). Измерения проводились в 8-10 радиальных рядах каждого из изученных годичных приростов (Антонова, Перевозникова, Стасова, 1999). На поперечных срезах также определяли количество клеток ранней и поздней древесины. Для приведения разного числа клеток в годичном слое модельных деревьев к единому было проведено нормирование.
Результаты и обсуждение
На рисунке 2 совмещенно представлена характеристика трахеид по площади и годичного радиального прироста. Следствием дефолиации является снижение радиального прироста, переход от ранней древесины к поздней становится более резким. При этом степень снижения положительно коррелирует со степенью уничтожения фотосинтезирующего аппарата. Максимальная потеря прироста характерна для активно растущих деревьев (в 5 и более раз).

|
а) |

|
в) |
Рисунок 1- Поперечный срез древесины сосны обыкновенной (а- в период дефолиации; в- контроль)
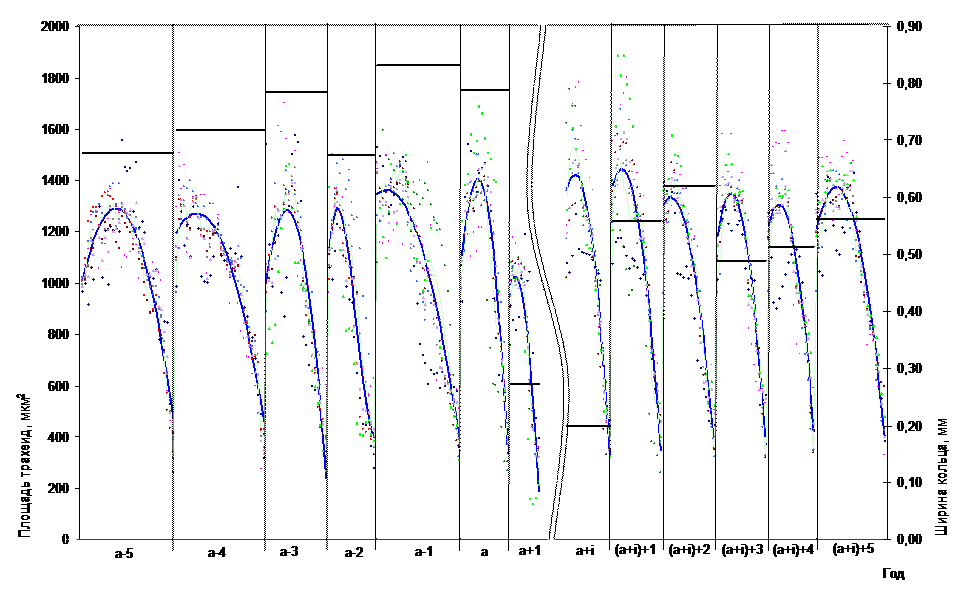
Рис. 2 Нормированная трахеидограмма и динамика радиального прироста (а - дефолиация; а+i - год начала интенсивного восстановления)
Ширину годичного прироста древесины определяют радиальные диаметры трахеид и их количество в радиальном ряду (Антонова, Шебеко, 1985). И.А. Терсков и др. (1978) утверждают, что изменение прироста происходит в основном за счет изменения числа клеток, а не их размеров.
Закономерности распределения морфометрических характеристик трахеид вдоль радиального ряда клеток интересны при оценке результатов физиологических и биохимических процессов в определенные периоды вегетационного сезона при воздействии различных экзогенных факторов. Наиболее крупные клетки образуются в начале вегетационного периода и в дальнейшем отмечается постепенное, а к границе ранней и поздней древесины значительное снижение радиальных размеров трахеид (Антонова 1999). Внешние размеры трахеид определяются степенью развития первичной клеточной стенки. В фазу роста растяжением длина трахеальных производных камбия увеличивается по сравнению с исходной длиной веретеновидных инициалей в среднем на 20 % (Крамер, Козловский, 1983).
В связи с тем, что процессы продукции клеток, рост растяжением и утолщения клеточных стенок, с одной стороны, находятся под контролем внутренних факторов дифференциации ксилемы в сезонном цикле, с другой - испытывают прямое и косвенное влияние внешних условий на эти процессы связь ширины годичного кольца с размером клеток характеризуется сложными зависимостями (Ваганов, Шашкин, 2000). Так, для относительно широких (с приростом более 0,5-0,6 мм) колец лиственницы и ели на полярном пределе их распространения зависимость между средними размерами клеток в ранней и поздней древесине от ширины годичного кольца практически отсутствует. Для сосны, произрастающей в подзоне средней тайги, увеличение радиальных размеров трахеид с ростом ширины годичного кольца прекращается на уровне «очень широких»: 2,16-3,37 мм (Свидерская, 1999). Причина заключается в возможном существовании максимального размера люмены, превышение которого ведет к нарушению проводящих функций.
Установлена умеренная связь между радиальным приростом и площадью ранних трахеид в периоды до и после дефолиации, а также в контроле (табл. 1). Однако, при включении в анализ всего изученного периода корреляция исчезает. Причина заключается в нарушении существующих закономерностей как ответная реакция древесных на дефолиацию. В контроле корреляция между данными показателями высокая (0,73).
Таблица 1 Зависимость между текущим радиальным приростом и площадью составляющих его трахеид
Древесина |
До дефолиации |
Дефолиация и восстановление |
После дефолиации |
Весь период |
Контроль |
а-1(n=5) |
от а+1до а+i |
от а до а+i |
(а+i)+1(n=5) |
r |
p |
r |
p |
r |
p |
r |
p |
r |
p |
r |
p |
Ранняя |
0,57 |
0,0005 |
-0,41 |
>0,05 |
-0,02 |
0,89 |
0,61 |
0,001 |
0,11 |
>0,05 |
0,73 |
0,0001 |
Поздняя |
0,40 |
0,02 |
-0,63 |
>0,05 |
-0,06 |
0,71 |
0,31 |
0,2 |
0,13 |
>0,05 |
0,58 |
0,0001 |
На рисунке 3 изображено изменение средних значений площади ранних и поздних трахеид. В год дефолиации отсутствует какое-либо снижение радиального прироста и площади клеток. Причина заключается в позднем объедании хвои гусеницами СШ. Отмечается лишь некоторое снижение доли поздней древесины. На следующий год резко (более чем в два раза) уменьшается ширина годичного кольца и площадь образующих его клеток. Средняя площадь ранних трахеид снижается на 21 %; поздних - на 11 %. Во время вспышки массового размножения СШ возможно неоднократное объедание одного и того же дерева. Поэтому период восстановления выбранной для исследования совокупности деревьев растягивается на период от 2 до 5 лет. При этом в период восстановления обнаружена интересная закономерность, ранее никем не отмечаемая. Увеличению радиального прироста предшествует рост площади трахеид (на 35 % - как ранних, так и поздних). В таблице 2 приведены коэффициенты корреляции между ?шгк=ШГК(а+i)+1 –ШГК(а+1) (разница между шириной годичного кольца в первый год интенсивного восстановления прироста и предыдущего) и ?S =S(а+i) - S(а+1) (разница между площадью трахеид в первый год интенсивного восстановления прироста и в первый после дефолиации год снижения прироста).
В другие периоды (до дефолиации, после дефолиации) и в контроле для расчета взяты участки ксилемы с существенным увеличением прироста. Корреляционный анализ подтвердил высокую связь между увеличением ширины годичного кольца и предшествующим ростом площади трахеид только для периода интенсивного восстановления прироста (0,81, р=0,005). В другие периоды связь либо отсутствует, либо не достоверна (р>0,05). Взятые для анализа периоды до дефолиации и после дефолиации не имеют столь большого снижения прироста, как во время вспышки массового размножения вредителя. Поэтому в контрольных кернах был выбран период значительного снижения радиального прироста (сопоставимого с дефолиируемыми моделями), в основе которого лежат климатические и ценотические факторы. Указанная зависимость не найдена, что свидетельствует о специфичности данного параметра для идентификации вспышки массового размножения СШ.
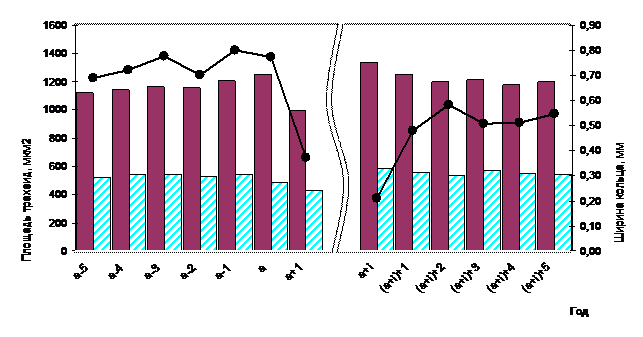
Рис. 3 - Динамика радиального прироста и средних значений площади ранних и поздних трахеид (а - дефолиация; а+i - год начала интенсивного восстановления)
Таблица 2 Зависимость между увеличением ширины годичного кольца и предшествующим ростом площади трахеид
Древесина |
До дефолиации |
Восстановление |
После дефолиации |
Контроль |
r |
p |
r |
p |
r |
p |
r |
p |
Ранняя |
-0,38 |
>0,05 |
0,81 |
0,005 |
0,05 |
>0,05 |
0,16 |
>0,05 |
Поздняя |
-0,56 |
>0,05 |
0,81 |
0,005 |
0,05 |
>0,05 |
0,60 |
>0,05 |
Отмеченная закономерность имеет в своей основе как минимум три причины:
1- улучшение условий роста в результате гибели рядом растущих деревьев (менее устойчивых к дефолиации пихты, ели, кедра). Как следствие - активизация всех физиолого-биохимических процессов, ведущих к увеличению количества и размеров трахеид, радиального прироста;
2 - увеличение количества влаги в хвое, лубе. Не вызывает сомнения рост количества влаги в почве (часто отмечается заболачивание) из-за гибели большого количества деревьев. При этом установлено, что для сосны из влажных условий местопроизрастания характерны более крупные клетки (Vysotskaya, Vaganov, 1989). С возрастанием дефицита влаги уменьшается средний размер клеток как в ранней, так и поздней древесине (Ваганов, Шашкин, 2000). Согласно теории клеточного роста, тургорное давление является необходимым условием роста (Kozlovski, 1968). На этом основано заключение, что конечный размер трахеид определяется условиями в период растяжения клеток (Ваганов, Шашкин, 2000). При дефолиации значительно и резко уменьшается отношение объема хвои к массе корневой системы. Как реакция на частичную дефолиацию, у деревьев резко возрастает интенсивность транспирации (у кедра - 182-320 %; у лиственницы - до 220 %, вторичная хвоя -до 255 %) (Плешанов, 1982). Повышение интенсивности транспирации при частичной дефолиации крон отмечено также у сосны и ели (Пиндюра, Чубук, 1975). Установлено, что с увеличением интенсивности транспирации образуется больше крупнопросветных тонкостенных трахеид у хвойных (Крамер, Козловский, 1983; Хари и др., 1985; Кайбияйнен и др., 1986). Остающаяся первичная и отрастающая в год дефолиации вторичная хвоя лиственницы отличается повышенной обводненностью (Васильева, Плешанов, 1975; Гирс, 1982). При достаточной почвенной влагообеспеченности повышение обводненности луба, особенно в прикомлевой части ствола, установлено при частичной дефолиации кедра сибирским шелкопрядом (Пиндюра, Плешанов, 1975). Некоторое снижение потенциала влаги в лубе дуба, поврежденного зеленой дубовой листоверткой, установлено для периода после восстановления листвы (Иерусалимов, 2004). Вероятнее всего, достоверное уменьшение потенциала влаги связано не с нарушением водного обмена, но с обеднением луба у них углеводами, которое обычно наблюдается у деревьев, перенесших дефолиацию и последующее восстановление листвы. Причина высокой обводненности луба заключается как в резкой потере органов транспирации (вся поднятая влага остается в стволе), так и в некоторой инерционности неповрежденной корневой системы, продолжающей активно поглощать воду. Е.Н. Иерусалимов (2004) отмечает, что у сосны, поврежденной сосновой пяденицей, поднятие пасоки замедляется, но не прекращается даже при полном повреждении крон (оставаясь на уровне 30 % от контроля).
3- первоочередное восстановление органов, определяющих дальнейший рост и развитие. А.С. Рожков и др. (1991) указывают на то, что радиальный прирост ствола нормализуется раньше линейного прироста побегов. А.С. Плешанов (1982) отмечает, что у кедра после весеннего повреждения длина хвои, формирующейся в текущем году, оказывается близкой к норме, а на следующий год превышает размеры хвои здоровых деревьев. Дефолиация деревьев приводит к перераспределению пластических веществ. Они используются преимущественно в частях растения, имеющих наибольшее значение для восстановления кроны и нормализации процессов фотосинтеза (Плешанов, 1982). Соотношение важности основных функций древесины (механическая и проведение водного тока) после дефолиации несколько изменяется. Уменьшение массы кроны снижает важность механической функции и возрастает роль трахеид, проводящих воду и минеральные вещества. Недостаток пластических веществ сопровождается их перераспределением - образуются крупные тонкостенные клетки. Простейший расчет показывает, что для образования 1 см2 люмен у дефолиированных деревьев, имеющих крупные трахеиды, требуется до 25 % меньше веществ, чем для деревьев, у которых клетки ранней древесины имеют обычные размеры.
Для некоторых деревьев (7с) даже на следующий после дефолиации год установлено значительное (+40%) увеличение площади ранних трахеид. Это является дополнительным подтверждением весомости влияния увеличения жизненного пространства и количества влаги.
На рисунке 4 приведены результаты анатомических исследований годичных слоев древесины сосны, растущей в условиях гарантированного отсутствия объедания хвои СШ (контроль). Выбран период значительного снижения радиального прироста, в основе которого - климатические и ценотические факторы.
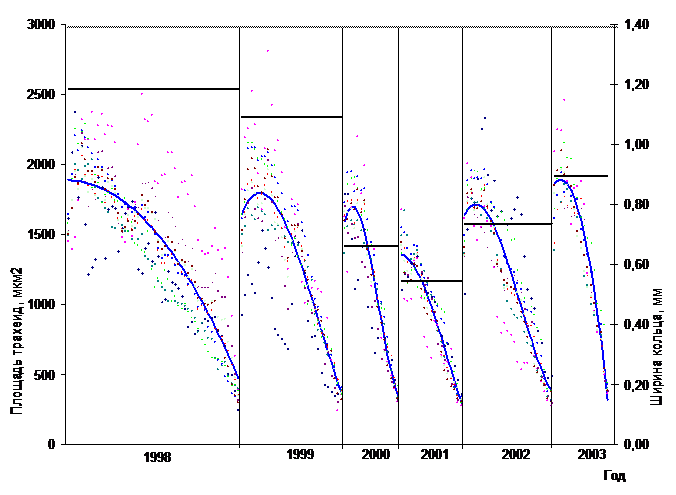
Рис. 4 Нормированная трахеидограмма и динамика радиального прироста (контроль)
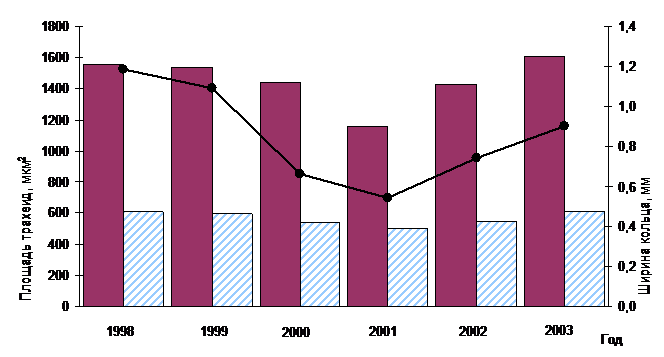
Рис. 5- Динамика радиального прироста и средних значений площади ранних и поздних трахеид (контроль)
В контрольных условиях уменьшение ширины годичного кольца сопровождается редукцией количества и площади трахеид как ранней, так и поздней древесины. При этом восстановление прироста, в отличие от «шелкопрядников», происходит одновременно с ростом площади клеток.
Выводы
- Следствием дефолиации является снижение радиального прироста, переход от ранней древесины к поздней становится более резким.
- Установлена умеренная связь между радиальным приростом и площадью ранних трахеид в периоды до и после дефолиации, а также в контроле (0,57 – 0,73). Однако, при включении в анализ всего изученного периода - корреляция исчезает. Причина заключается в нарушении существующих закономерностей как ответная реакция древесных на дефолиацию.
- В год дефолиации отсутствует какое-либо снижение радиального прироста и площади клеток. Причина заключается в позднем объедании хвои гусеницами СШ. Отмечается лишь некоторое снижение доли поздней древесины. На следующий год резко (более чем в два раза) уменьшается ширина годичного кольца и площадь образующих его клеток. Средняя площадь ранних трахеид снижается на 21 %; поздних - на 11 %. Уменьшается толщина клеточных стенок, более значительно - в зоне поздней древесины.
- Увеличению радиального прироста в период активного восстановления предшествует рост площади трахеид (на 35 % - как ранних, так и поздних). Коэффициент корреляции равен 0,81 (р<0,005). Отмеченная закономерность имеет в своей основе как минимум три причины: а- улучшение условий роста; в - увеличение количества влаги в хвое и лубе; с- первоочередное восстановление органов, определяющих дальнейшие рост и развитие.
Литература:
- Антонова Г.Ф. Рост клеток хвойных. - Новосибирск: Сибирская издательская фирма «Наука» РАН, 1999. – 232 с.
- Антонова Г.Ф., Перевозникова В.Д., Стасова В.В. Влияние условий произрастания на структуру годичного слоя древесины и продуктивность сосны обыкновенной. // Лесоведение. – 1999. - № 9. – с.45-53.
- Антонова Г.Ф., Шебеко В.В. Формирование ксилемы хвойных. 3. Динамика развития трахеид в зонах дифференциации // Лесоведение. -1985. -№ 5. С. 71-74.
- Арсеньева Т. В., Чавчавадзе Е. С. Эколого-анатомические аспекты изменчивости древесины сосновых из промышленных районов европейского Севера. — СПб.: Наука, 2001. — 109 с.
- Ваганов Е.А., Терсков И.А. Анализ роста дерева по структуре годичных колец. — Новосибирск: Наука. Сиб. отд-ние, 1977. — 93 с.
- Ваганов Е.А., Шашкин А.В. Рост и структура годичных колец хвойных. - Новосибирск: Наука, 2000. — 232 с.
- Васильева Т. Г., Плешанов А. С. Физиология ослабления и отмирания лиственницы в очагах пяденицы Якобсона //Влияние антропогенных и природных факторов на хвойные деревья. Иркутск , 1975, 179-214.
- Гирс Г.И. Физиология ослабленного дерева.- Новосибирск: Наука, 1982. -256 с.
- Дудин В.А. Проблема использования и восстановления шелкопрядников в Томской области //Труды по лесному хозяйству Сибири, вып. 4. -Новосибирск, 1958. - С. 262 - 268.
- Иерусалимов Е.Н. Зоогенная дефолиация и лесное сообщество. –М.: Товарищество научных изданий КМК, 2004. -263 с.
- Исаев А.С., Хлебопрос Р.Г., Недорезов Л.В., Кондаков Ю.П., Киселев В.В., Суховольский В.Г. Популяционная динамика лесных насекомых. -М.: Наука, 2001. – 374 с.
- Кайбияйнен Л.К., Хари П., Сазонова Т.А. Сбалансированность системы водного транспорта у сосны обыкновенной. 3. Площадь проводящей ксилемы и масса хвои //Лесоведение. -1986. -№. 1. -С. 445-451.
- Коломиец Н. Г., Майер Э. И. Важнейшие вредители лесов Томской области и меры борьбы с ними. –Томск: Книж. изд-во, 1963.
- Коломиец Н.Г. Сибирский шелкопряд и его роль в хвойных лесах Западной Сибири // Труды по лесному хозяйству Сибири. вып. 7. - Издательство СО АН СССР, Новосибирск, 1962. - С. 137 - 161.
- Косиченко Н. Е. Влияние генотипа — среды — на формирование микроструктуры стебля и диагностика технических свойств роста и устойчивости древесных растений: Дис. ... д-ра биол. наук. Воронеж, 1999. - 298 с.
- Крамер П.Д., Козловский Т.Т. Физиология древесных растений: Пер. с англ. — М.: Лесн. пром-сть, 1983. — 464 с.
- Крылов Г.В. Леса Сибири и Дальнего Востока, их лесорастительное районирование. –М.-Л.: Гослесбумиздат, 1960. -156 с.
- Крылов Г.В., Таланцев Н.К., Куликов М.И., Демиденко В.П., Храмова Н.Ф. Лесовозобновительные процессы в лесах таежной зоны Западной Сибири //Возобновление леса. Научные труды ВАСХНИЛ. -М.: Колос, 1975. - С. 252 - 272.
- Лир Х., Польстер Г., Фидлер Г.И. Физиология древесных растений. – М.: Лесн. Пром-сть, 1974. – 421 с.
- Пиндюра Е С., Чубук С. А. О транспирации дефолиированных хвойных деревьев.— Информ. матер. СИФИБРа, 1975, вып. 13. -С. 19-20.
- Пиндюра Е. С., Плешанов А. С. Морфометрические реакции хвойных на дефолиацию //Влияние антропогенных и природных факторов на хвойные деревья. Иркутск, 1975. -С. 159— 178.
- Плешанов А.С. Насекомые – дефолианты лиственничных лесов Восточной Сибири. – Новосибирск: Наука, 1982. -209 с.
- Рожков А.С., Хлиманкова Е.С., Степанчук Е.С. Восстановительные процессы у хвойных при дефолиации. –Новосибирск: Наука, 1991. -88 с.
- Павлов И.Н., Мочалов А.А., Агеев А.А., Ничепорчук В.В., Россинин В.В. Биометрия клетки древесины. Свид. 2005612253 РФ. / (RU. - № 2005611740; Заявл. 11.07.05; Опубл. 02.09.05, Бюл. № 9).
- Свидерская И.В. Гистометрический анализ закономерностей сезонного формирования древесины хвойных: Автореф. дис. канд. биол. наук: 06.03.01. Красноярск. 1999. - 22с.
- Свидерская И.В., Пальникова Е.Н. Радиальный рост сосны в связи с дефолиацией сосновой пяденицей // Лесоведение. - 2003. - № 5. - С. 44-53.
- Терсков И. В., Ваганов Е.А., Зырянов Г.И., Филимонов В.С. Анализ роста дерева с помощью кривых распределения клеток по размерам внутри годичных колец // Изв. СО АН СССР. Сер. биол. Наук. – 1978. – Вып. 3(15). – С. 26-36.
- Фалалеев Э.Н. Естественное возобновление пихтовых лесов, поврежденных сибирским шелкопрядом. // Лесной журнал. - 1960. - № 2. - С. 35 - 37.
- Федоренко С.И. О выпадении годичных колец в процессе формирования ксилемы березы бородавчатой // Исслед. лесов Урала : Матер. науч. чтений, посвящ. памяти Б.П. Колесникова. - Екатеринбург, 1997. - С. 69-71.
- Фуряев В.В. Шелкопрядники тайги и их выжигание. -М.: Наука, 1966. - 92 с.
- Хари П., Кайбияйнен Л. К., Сазонова Т. А., Мякеля А. Сбалансированность системы водного транспорта у сосны обыкновенной. 2. Активная ксилема // Лесоведение. -1985. -№ 5. -С. 74—75.
- Чавчавадзе Е.С. Древесина хвойных. - Л.: Наука. Ленингр. отд-ние, 1979. — 192 с.
- Filion L., Cournoyer L. Variation in wood structure of eastern larch sawfly in subarctic Quebec, Canada // Can. J. For. Res. - 1995. - Vol. 25. - P. 1263- 1268.
- Hogg E.H., Hart M., Lieffers V.J. White tree rings formed in trembling aspen saplings following experimental defoliation // Can. J. Forest Res. - 2002. - 32, № 11. - Р. 1929-1934.
- Koslowski T.T. Tree physiology and tree pests // J. Forestry. -1969. -Vol.67. -P. 118-122.
- Schweingruber F.H. Auswirkungen des Larchenwicklerbefalls auf die Janrringesstructur der Larche; Ergebnisse einer Jahrringanalyse mit rentgendensitometrische Methoden. // Schweiz. Z. Forstwesen. — 1979.
- Seija A., Riikka P., Jari O., Pia M/, Pekka S., ElinaV. Effects of defoliation on growth, biomass allocation, and wood properties of Betula pendula clones grown at different nutrient levels // Can. J. Forest Res. - 2002. - 32, № 3. - Р. 498-508.
- Vysotskaya L.G., Vaganov E.A. Components of the variability of radial cell size in tree rings of conifers // IAWA Bull. - 1989. - Vol. 10, № 4
|


