|
"Хвойные бореальной зоны" 2009г., №1, с. 48-57
Влияние лесорастительных условий на устойчивость сосняков Минусинской впадины к корневым патогенам
И.Н. Павлов, П.В. Губарев, О.А. Барабанова, А.А. Агеев, Ю.А. Орлов, А.И. Лобанов
ГОУ ВПО «Сибирский государственный технологический университет»
660049 Красноярск, пр. Мира, 82; e-mail: forester24@mail.ru
В работе приведены результаты исследования влияния изменения климата, антропогенного воздействия, эдафических условий, биологической продуктивности, внутриценотической конкуренции на устойчивость сосновых лесов к корневым гнилям, вызываемым Heterobasidionannosum (Fr.) Bref s. str. и Armillariamelleas.l.
Ключевые слова: Heterobasidionannosum (Fr.) Bref s. str., Armillariamelleas.l., Минусинская впадина
The results of climate change study, anthropogenic influence, edaphic conditions, bioproductivity, study of intracenosis competition in resistance of pine forests to root rot caused by Heterobasidion annosum (Fr.) Bref s. str. and Armillaria mellea s.l. are given in this work.
Key words: Heterobasidion annosum (Fr.) Bref s. str. и Armillaria mellea s.l., Minusinsk depression
Введение
Ухудшение санитарного состояния лесов бореальной зоны привлекает все большее внимание исследователей и общественности. Неблагоприятное изменение климата, интенсификация антропогенного воздействия, межрегиональное перемещение болезней и вредителей являются основными причинами современной дигрессии лесов (Nuorteva, 1997; Donaubauer, 1998; Tagliaferro, 1998; Rigling et al., 1999; Vertui, Bruhn et al., 2000; Hogg et al., 2002; Hogg, Brandt, Kochtubajda, 2002; Woodward, 2006; Алексеев и др., 1999; Стороженко, 2001; Hopper, Sivasithamparam, 2005; Павлов, 2007).
Конечно, разрушительные процессы в лесах существовали всегда и носили как быстрый характер (вспышки массового размножения сибирского шелкопряда, пожары, ветровал, и пр.), так и медленный (сукцессии по типу заболачивание — разболачивание или остепнение — залесение). В основе указанных процессов – климатические изменения. Тем не менее, всегда оставались лесные массивы, находящиеся в стабильном состоянии и в равновесии с климатическими условиями неопределенно долгое время (региональный климакс). Исследование лесных экосистем, находящихся в состоянии регионального (климатического) или эдафического климакса, особенно в условиях продолжающегося катастрофического антропогенного воздействия на них, представляет значительный интерес. Исключительно важно знать их закономерности роста, развития, строения для дальнейшего принятия хозяйственных решений при восстановлении нарушенных ранее ценозов.
Устойчивость к возбудителям корневых гнилей чрезвычайно важна для успешного роста большинства высших растений. Механизмы, обеспечивающие их устойчивость разнообразны. Некоторые из них достаточно легко воспроизводимы. Так, одним из параметров строения, способствующего высокой устойчивости, является пространственная изоляция растений. Например, распространение грибов рода Pythium, поражающих и губящих проростки многих высших растений, зависит от того, насколько близко друг от друга расположены проростки (рис. 1). Как в садовых, так и в лесных насаждениях, массовое заражение растений этим паразитом предотвращается за счет разреженных посевов. Чрезвычайно простой, но достаточно эффективный способ. Наряду с пространственной изоляцией, за счет большей площади питания обеспечивается более высокая биологическая устойчивость растений.
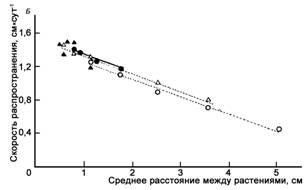
Рисунок 1 – Скорость распространения патогенного гриба Pythium irregulare в популяциях кресс-салата (Lepidium sativum) от зараженных проростков, помещенных у края посевов (по Burdon, Chilvers, 1975)
Из этого следует, что при искусственном лесовосстановлении при наличии инфекции корневых патогенов в почве целесообразно создание смешанных культур из разных видов или из высоко полиморфных популяций (в случае если только одна древесная порода является единственно возможной для произрастания). При этом эффективная плотность каждого вида (генотипа) снижается — она «разбавляется» другими. В смешанной культуре каждая устойчивая к заболеванию форма может служить барьером, сдерживающим распространение инфекции среди восприимчивых особей. К сожалению, при создании искусственных лесных насаждений и использовании семян от ограниченного числа деревьев, происходит снижение полиморфизма популяций (Шигапов, 1993).
Биологические и экологические свойства корневых патогенов (Armillariamellea s.l.; Heterobasidionannosum s.l.) определяют их достаточно высокую отзывчивость на любые изменения климата. Роль паразитов, хищников и болезней наименее существенна в формировании структуры сообществ, функционирующих в относительно суровых, изменчивых или непредсказуемых условиях (Connell, 1975). Но, по мере изменения условий произрастания к оптимальным для патогенных организмов, следует ожидать роста их вредоносного воздействия. Возможно, из-за отсутствия естественных врагов и антагонистов, даже в больших масштабах.
Программа, методика и объекты
Ленточные боры Минусинской котловины из сосновых лесов наиболее удалены к югу и являются интразональными образованиями. Располагаясь среди степей, на значительном удалении от морей и океанов, они испытывают весь комплекс неблагоприятных климатических воздействий (большие годовые амплитуды колебания температур, малое количество осадков, сухость воздуха). Отмечаемое
глобальное увеличение температуры приземного слоя воздуха (Изменение климата: физическая научная основа, 2007) в большей степени проявляется на границе природных зон, например, в экотоне лес-степь, где температура и, особенно, осадки являются лимитирующими факторами. Поэтому именно в этих условиях произрастания с изменением лесорастительных условий в первую очередь и следует ожидать активизации воздействия корневых патогенов. Лесорастительные условия включают весь комплекс климатических, орографических, гидрологических и почвенных факторов, определяющих условия роста лесной растительности и динамику леса. Естественно, лесорастительные условия определяют и устойчивость древесных растений к неблагоприятным воздействиям как биотического, так и абиотического происхождения.
Наряду с климатом, на устойчивость древесных растений к возбудителям корневых гнилей и развитие заболевания чрезвычайно значимое влияние оказывают почвенные условия. Однако, исследование влияния почвенного плодородия на патогенность Heterobasidionannosum s.l. и Armillaria mellea s.l. в естественных условиях, а так же опыты с применением минеральных удобрений показали неоднозначные результаты (Федоров, 1984; Негруцкий, 1986, Shaw, Kile, 1991; Heterobasidionannosum, 1998).
Исследуемые нами сосняки по степени увеличения плодородия и увлажнения образуют ряд: мертвопокровный ® бруснично- зеленомошный ® разнотравно-зеленомошный ® папоротниковый (табл. 1).
Таблица 1 – Характеристика постоянных пробных площадей
№ ППП |
Состав |
Возраст |
Бонитет |
Тип леса |
Почвы |
|
1 |
10С |
130 |
III |
Сосняк мертвопокровный |
Дерново-боровая примитивная супесчаная, сухая |
|
2 |
10С |
100 |
III |
Сосняк бруснично- зеленомошный |
Дерново-боровая супесчаная |
|
3 |
10С едБОc |
90 |
II |
Сосняк разнотравно-зеленомошный |
Серая лесная легкосуглинистая |
|
4 |
10С едЕ |
100 |
Ia |
Сосняк папоротниковый |
Темно-серая лесная среднесуглинистая, свежая |
|
При закладке постоянных пробных площадей и их обследовании использовались общепринятые в лесоведении, дендрохронологии, лесной таксации, почвоведении методические подходы и положения (Уткин, 1975; Кузьмичев, 1977; Почвы СССР, 1979; Емшанов, 1999; Ваганов, Шашкин, 2000). Выделение видов грибов, входящих в комплекс A. mellea s.l., проведено по классификации К. Корхонена (1978, 2004). Принадлежность корневой губки к H. annosum sensu stricto, вызвавшей значительное куртинное усыхание сосновых древостоев в Минусинских ленточных борах, установлена в результате тест-скрещивания, проведенного К. Корхоненом.
обсуждение и анализ результатов
В результате продолжающихся исследований изменения глобального климата подтвержден устойчивый рост температуры приземного слоя воздуха (Изменение климата: физическая научная основа, 2007). Обновленный 100-летний тренд за 1906–2005 годы, составил 0,74 0C ± 0,18 0C и превышает 100-летний тренд потепления (1901–2000 годы), отмеченный на момент выпуска предыдущей версии доклада Межправительственной группы экспертов по изменению климата, равный 0,6 0C ± 0,2 0C. В целом, темпы потепления за последние 50 лет (0,13 0C ± 0,03 0C за десятилетие), почти вдвое выше, чем за последние 100 лет.
Увеличение среднегодовой температуры приземного слоя воздуха за период 2001-2008 гг. относительно 1961-1990 гг. составило + 0,43 0C (рис.2). Это значительно теплее периода 1991-2000 гг. (рост составил +0,19 0C).
Годовое выпадение осадков на суше в течение 20 века возрастало в средних и высоких широтах Северного полушария (примерно 0,5-1 % в десятилетие), за исключением Восточной Азии (Technical Summary Working Group I, 2001). В свою очередь проведенная реконструкция климата за последние 450 лет по образцам древесины Larixsibirica, Pinussibiricaи P. sylvestris, полученных из горных лесов Монголии, расположенных на высоте от 900 до 2440 м над уровнем моря показала, что сейчас климат данного региона значительно теплее и влажнее, чем прежде (Jacoby et al., 1999).
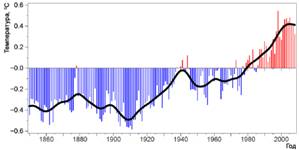
Рисунок 2 – Среднегодовая температура у поверхности Земли ( www.cru.uea.ac.uk)
В начале 21 века существующие тренды сохранились (Изменение климата: физическая научная основа, 2007). Существенно возросшее количество осадков наблюдалось в восточных частях Северной и Южной Америки, северной части Европы, северной и центральной частях Азии. Засушливость наблюдалась в Сахеле, Средиземноморье, южной Африке и частях южной Азии. В дальнейшем на планете так же ожидается увеличение количества осадков на высоких широтах, уменьшение более вероятно в большинстве субтропических районов суши (до 20 % к 2100 г.)
В последние десятилетия (1976 – 2007 гг.) в Европейской части России, Приамурье и Приморье происходит уменьшение летних осадков. В то же время в Средней Сибири – устойчивое увеличение. Коэффициент линейного тренда составляет +2,08 (максимальное значение для регионов России) (Обзор состояния …, 2007).
Для экосистем бореальных лесов предполагается, что влияние климатических изменений будет опосредовано в изменениях частоты пожаров, вспышек массового размножения вредителей, динамики прогалин (Bergeron et al., 1998). Потенциальная опасность для сосновых лесов от сосновой пяденицы в ряде районов Центральной Сибири возникает уже при повышении температуры на 2-4 °С (Исаев и др., 1999).
Связь корневых патогенов с изменением температуры и влажности среды достаточно велика (Shaw, Kile, 1991; Heterobasidion annosum, 1998). Оптимальными температурами для роста мицелия опенка являются 19-25 0С (Соколов, 1964; Shaw, Kile, 1991) (рис. 3). При культивировании на агаре оптимальная температура для роста выше и достигает 28 0С (Rishbeth, 1978).
Для роста и развития ризоморф температурный оптимум понижается и ограничивается 17,6-24,2 0С (Соколов, 1964), 22 0С (Rishbeth, 1968). Ниже 11 0С и выше 25 0С их рост очень слабый, а при температуре ниже 5 0С и выше 35 0С рост полностью прекращается. В идентичных условиях ризоморфы растут в 13 раз быстрее мицелия (Rishbeth, 1968). Скорость распространения ризоморф в верхнем слое почвы может превышать один метр в год (Redfern, 1973).
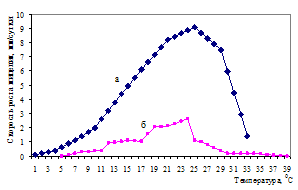
Рисунок 3 – Влияние температуры на рост мицелия корневой губки (а) и опенка (б) (Соколов, 1964; Негруцкий, 1986)
Более северные европейские виды A. borealis, A. cepistipes, A. ostoyaeимеют более низкий температурный оптимум, чем южные (из низких широт) видыA. gallicaи A. mellea(Guillaumin at al., 1989). Можно сделать предположение о большей устойчивости сибирских штаммов опенка к низким температурам. Но это требует дополнительных исследований.
Корневая губка менее чувствительна к воздействию низких температур. Рост мицелия начинается при температуре 0-2 0C, оптимальным является 22-28 0C (несколько выше, чем у опенка); рост прекращается при температуре 32-37 0C; грибы погибают в течение двух часов при 38-45 0C (Courtois, 1972, 1973; Негруцкий, 1973, 1986; Schwantes et at, 1976). При температуре среды 12 0C рост мицелия составляет только 50 % от роста при оптимальной температуре (Roll-Hansen, 1940), при 8 0C – около 21 % (Cowling, Kelman, 1964).
Мицелий корневой губки способен переносить чрезвычайно низкие температуры (до -270 0С) без потери жизнеспособности и поэтому причина гибели, очевидно, заключается не в абсолютном значении низких температур, а в длительности периода их воздействия (Негруцкий, 1986). При продолжительном действии низких температур питательная среда становится недоступной для усвоения, и гриб погибает после израсходования запасных питательных веществ.
В отличие от опенка, для корневой губки из холодных и теплых мест произрастания до настоящего времени не найдено больших региональных отличий по отношению к температурным условиям роста (Heterobasidion annosum, 1998). С нашей точки зрения это утверждение требует дополнительных исследований. Не менее важным для корневой губки является и влажность субстрата. Нижний порог влажности, необходимой для развития гриба, находится в пределах 20 % (Негруцкий, 1973).
Исходя из выше приведенных фактов можно заключить, что даже незначительные изменения климата окажут определяющее влияние на состояние хвойных ценозов, особенно произрастающих в экотоне лес-степь.
Минусинская котловина выделяется особо благоприятными для роста сельскохозяйственных культур климатическими условиями. Некоторые из этих условий также положительны и для насекомых, и для патогенных организмов. В дальнейшем с увеличением количества осадков и потеплением климата следует ожидать новых вспышек массового размножения насекомых, активизации болезней.
По данным Cредне-Сибирского межрегионального территориального управления по гидрометеорологии и мониторингу окружающей среды, для исследуемой территории характерен рост температуры приземного слоя воздуха, наиболее существенный в последние два десятилетия (рис. 4). Более теплая зима (с 1915 г. по
Все ветровальные деревья в большей или меньшей степени имели гниль, вызываемую Heterobasidionannosum (Fr.) Bref s. str. По степени поражения корней все деревья сосны были распределены на три группы: 1- без признаков поражения; 2- среднее; 3- сильное. На приведенной гистограмме показано влияние поражения корней на интенсивность ростовых процессов (рис. 9).
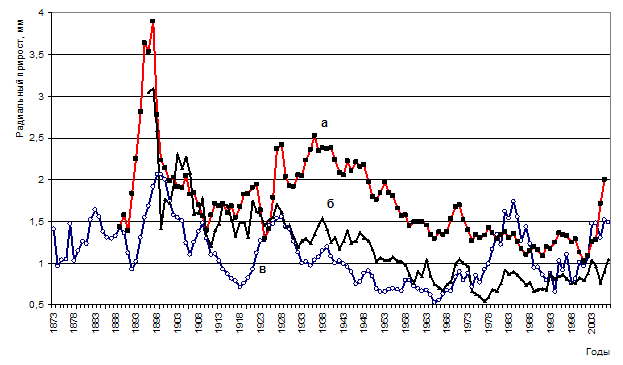
Рисунок 8 – Динамика радиального прироста в сосняках: мертвопокровном (а), зеленомошном (б), разнотравно-зеленомошном (в)
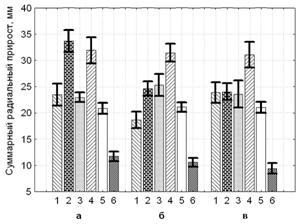
Рисунок 9 – Суммарный радиальный прирост ветровальных деревьев сосны обыкновенной (степень поражения корней Heterobasidionannosum (Fr.) Bref s. str.: а- без признаков поражения; б- среднее; в- сильное) по периодам (1 – 1873-1897; 2 – 1898-1918; 3 – 1919-1939; 4 – 1940-1979; 5 – 1980-1997; 6 – 1998-2007 г.г.)
Подтверждается закономерность, отмечаемая нами ранее (Павлов, 2007). Для устойчивых деревьев (без признаков поражения возбудителями корневых гнилей) характерно достаточно свободное произрастание в молодом возрасте, о чем и свидетельствует их более интенсивный рост. Они смогли избежать влияния «эффекта группового угнетения» (Погребняк, 1968; Романовский, 2002), который имеет место в условно-одновозрастном древостое (см. рис. 6) при достаточно равномерном размещении отдельных особей на площади после смыкания крон и вступления соснового древостоя в фазу жесткой конкуренции и активной дифференциации. Это подтверждает высокую опасность длительного нахождения соснового древостоя в условиях избыточной загущенности без активной дифференциации и своевременного отпада.
На второй пробной площади установлено единичное усыхание деревьев сосны как от воздействия Armillaria mellea s.l. так и Heterobasidionannosum (Fr.) Bref s. str. Насаждение расположено на несколько повышенном элементе рельефа (в сравнении с третьей ППП) на почве более легкого гранулометрического состава. И как следствие – значительно меньший радиальный прирост (см. рис. 8).
Наиболее неблагоприятное лесопатологическое состояние характерно для третьей постоянной пробной площади, где наибольшую опасность для деревьев сосны представляет опенок. Сухостой, образовавшийся за достаточно короткий период (до 10 лет) достигает 26 % от общего запаса стволовой древесины сосны (рис. 10).
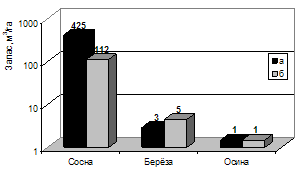
Рисунок 10 – Таксационная характеристика сосновых древостоев, имеющих очаговое поражение (а - живые; б - сухостой)
Веера мицелия опенка были обнаружены под корой усыхающих деревьев и свежего сухостоя. В местах предварительно проведенных раскопок корневых систем для стимулирования образования базидиом корневой губки (как предполагалось при постановке эксперимента) образовались плодовые тела Armillaria mellea s.l.. Базидиомы Heterobasidionannosum (Fr.) Bref s. str. были обнаружены только на верхней части исследуемого участка.
Распределение числа деревьев по диаметру является важным показателем, характеризующим состояние древостоя. Успешная аппроксимация распределения деревьев по диаметру в очагах корневых патогенов оказалась возможной только с использованием многокомпонентной смеси нормальных распределений (рис.11). Ряды распределения усохших деревьев имеют ярко выраженную положительную асимметрию.
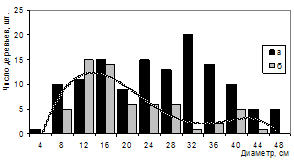
Рисунок 11 – Распределение числа живых (а) и усохших (б) деревьев сосны обыкновенной по диаметру (ППП 3)
Наличие сухостоя диаметром 16-44 см свидетельствует об интенсивном патогенном характере отпада. Первая составляющая отпада (в нижних ступенях толщины) образована патогенным воздействием корневых патогенов на деревья сосны, ослабленные внутриценотическим давлением.
В дальнейшем именно на их основе происходит усиление вирулентности и агрессивности опенка. Поэтому технологические решения, как при естественном возобновлении, так и при искусственном восстановлении должны быть направлены на предотвращение образования значительно угнетенного яруса деревьев. Например, за счет редкого размещения посадочного материала или своевременного проведения рубок ухода. Вторая составляющая отпада (в верхних ступенях толщины) образована действием опенка, как первичного патогена.
Двухвершинное распределение характерно и для живых деревьев. Причиной образования сложного строения живых деревьев является их возрастное строение (рис. 12) и патогенное воздействие корневых патогенов.
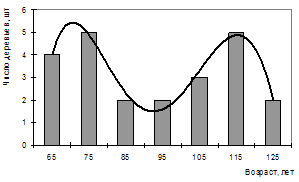
Рисунок 12 – Распределение числа деревьев сосны обыкновенной по возрасту (ППП 3)
Анализ радиального прироста живых и усохших деревьев в очагах корневых патогенов выявил существенные различия в динамике (рис. 13).
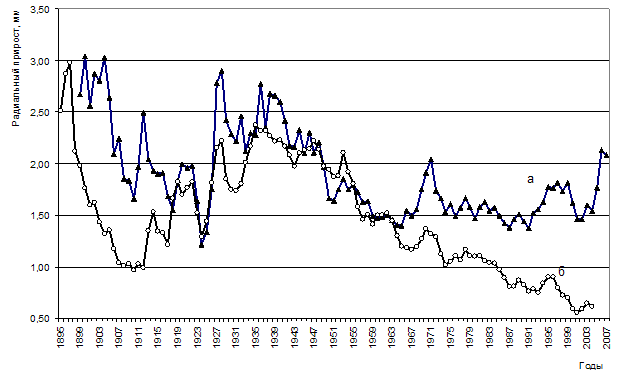
Рисунок 13 – Динамика радиального прироста живых (а) и погибших (б) деревьев в сосняке разнотравно-зеленомошном (ППП 3)
Заметные различия в радиальном приросте прослеживаются в периоды 1900-1911 гг. и 1929-1943 гг. Это периоды активизации внутриценотической конкуренции, начавшейся после смыкания крон. Живым в настоящее время деревьям удалось успешно пройти стадии дифференциации и своевременного отпада без значительного снижения своего прироста и жизнеспособности (естественно за счет отпада рядом растущих деревьев). С 60-х годов наблюдается значительное снижение радиального прироста у усохших в настоящее время деревьев. Возможно, наряду с ростом конкуренции за свет, воду, питательные вещества, именно в этот период начался процесс поражения корневых систем возбудителями корневых гнилей.
На четвертой пробной площади (сосняк папоротниковый на темно-серой лесной почве, мощный гумусовый горизонт, благоприятный гидрологический режим), расположенной на территории, прилегающей к пойме р. Лугавка, в наиболее оптимальных для сосны обыкновенной условиях произрастания, несмотря на присутствие опенка в подстилке, на мертвом подросте сосны и ели, отсутствуют усыхающие деревья I-IV классов Крафта. Симптомы поражения корневой губкой отсутствуют. На более высоких элементах рельефа прилегающего леса из корневых патогенов доминирует Heterobasidionannosum (Fr.) Bref s. str., ниже, в пойме р. Лугавка – Armillaria mellea s.l. (хотя и в том и другом случае были найдены плодовые тела и следы поражения обоими возбудителями корневых гнилей).
На темно-серых почвах Минусинских боров с непромывным типом водного режима, который определяется высокой влагоемкостью покровного суглинка, зависанием влаги осадков при переходе суглинка в связные супеси и пески, а также наличием (местами) на глубине 2—5 м подстилающего карбонатного суглинка и, наконец, экранирующим влиянием сезонной мерзлоты формируются высокопродуктивные сосняки I и II классов бонитета (Почвенные факторы …, 1976). В этих условиях элементы питания, выносимые осадками вглубь, остаются в корнеобитаемой толще и с помощью десукции возвращаются в верхнюю высокогумусную плодородную толщу.
Только на данном участке, благодаря расположению на припойменной террасе, древостой не испытывает недостатка влаги, даже в неблагоприятные засушливые периоды. Следует отметить, что периодически наступающие засухи с характерными для них длительным бездождьем, повышенными температурами, суховеями и пыльными бурями оказывают значительное влияние на состояние Минусинских боров. При выпадении осадков за май-июнь мене 80 мм водные запасы в верхней толще почвы падают почти до влажности завядания (Почвенные факторы …, 1976). Это ведет к резкому снижению веса закладываемой хвои, происходит массовый отпад старой хвои, усыхание подроста, снижение текущего прироста и биологической устойчивости.
Сочетание высокой продуктивности и устойчивости на ППП 4 представляет наибольший интерес для исследования. Насколько велики различия между двумя крайними по условиям произрастания насаждениями видно из графиков распределения по высоте и диаметру (рис. 14, 15).
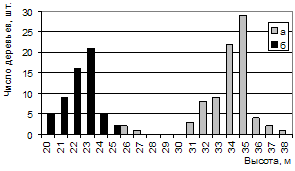
Рисунок 14 – Распределение числа деревьев сосны на ППП 4 (а) и ветровальных деревьев на ППП 1 (б) по высоте
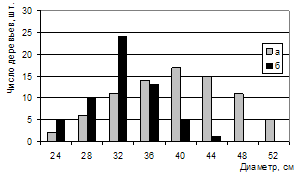
Рисунок 15 – Распределение числа деревьев сосны на ППП 4 (а) и ветровальных деревьев на ППП 1 (б) по диаметру
Причина устойчивости древостоя в данных условиях произрастания заключается в ряде сопряженных факторов:
- большую биологическую устойчивость деревьев обеспечивает глубоко развитая корневая система, оптимальный водный и питательный режим, хорошо развитая крона;
- высокая микробиологическая активность почв обеспечивает быстрое разложение опада и других растительных остатков. Это ухудшает условия сапротрофного питания корневых патогенов и снижает скорость их распространения в почве. В то же время в подверженных засухе древостоях (ППП 1, 2), наряду со снижением микробиологической активности почвы, повышенный отпад подроста (или значительное снижение биологической устойчивости) обеспечивает благоприятные условия для развития Heterobasidionannosum (Fr.) Bref s. str. и Armillaria mellea s.l.
- хорошо развитый живой напочвенный покров из крупнотравных видов растений исключает развитие зеленых мхов. В их отсутствие корневая шейка сосны обыкновенной менее подвержена поражению ризоморфами опенка (в слое мха, растительного опада кора сосны размягчается и становится легко проницаемой для ризоморф).
- припойменное расположение древостоя обеспечивает устойчивое обеспечение деревьев сосны влагой в любой засушливый период (чрезвычайно важно в условиях недостатка осадков, сухости воздуха, типичных для Минусинской котловины).
Выводы
- Тренд увеличения температуры приземного слоя воздуха на территории Минусинских боров за три последних десятилетия составил: по среднегодовой + 1,5 0С; по зимней + 2,8 0С; по температуре мая + 1,6 0С. За тот же период продолжительность теплого периода с температурой более 10 0С увеличилось на шесть дней. С 1936 г. по 2007 г. тренд увеличения максимальной скорости ветра при порыве составил 10 м/с.
- Указанные климатические изменения в условиях дефицита влаги формируют условия стресса для древесных растений Минусинских боров. В то же время увеличение продолжительности вегетационного периода, ранняя весна, более мягкие зимы благоприятны для корневых патогенов. Рост ветровой нагрузки наряду с вывалом деревьев увеличивает вероятность механического повреждения корней, что способствует распространению возбудителей корневых гнилей.
- В сухих олиготрофных условиях произрастания на супесчаных дерново-боровых примитивных почвах установлена приуроченность очагов усыхания к наличию прослоек суглинка среди песчаных отложений, что обеспечивает формирование оптимальной для корневой губки влажности (во влажный период - отвод избытка влаги благодаря песчаному обрамлению очагов; в засушливый период - её подъем (аккумуляция) по суглинистым элементам). За пределами суглинистых прослоек очаги усыхания обнаружены не были. Влияние климата нелинейно. С одной стороны быстрое прогревание песчаных почв способствует увеличению скорости распространения мицелия Heterobasidionannosum (Fr.) Bref s. str., с другой – сухость условий произрастания снижает её. Влияние Armillaria mellea s.l. ограничивается ростом на угнетенных деревьях сосны и березы в естественных понижениях между грив и не представляет значительной опасности.
- В свежих мезотрофных условиях произрастания на серой лесной легкосуглинистой почве установлено наиболее неблагоприятное санитарное состояние. Наибольшую опасность для деревьев сосны представляет Armillaria mellea s.l. Веера мицелия опенка были обнаружены под корой усыхающих деревьев и свежего сухостоя. Базидиомы Heterobasidionannosum (Fr.) Bref s. str. были обнаружены только на верхней части исследуемого участка.
- В свежих эвтрофных условиях произрастания на темно-серых лесных почвах высокая биологическая устойчивость и продуктивность древостоя сосны обеспечиваются глубоко развитой корневой системой; оптимальным водным и питательным режимом; хорошо развитой кроной; отсутствием толстой подстилки (за счет высокой микробиологической активность почв) – субстрата для распространения мицелия Heterobasidionannosum (Fr.) Bref s. str. и Armillaria mellea s.l.; отсутствием подушки из зеленых мхов, что обеспечивает естественное подсушивание корневой шейки сосны (в слое мха, растительного опада кора сосны размягчается и становится легко проницаемой для ризоморф опенка).
- Во влажных и сырых эвтрофных условиях произрастания наибольшую опасность для сосны обыкновенной, лиственницы сибирской, ели сибирской представляет Armillaria mellea s.l.. Базидиомы Heterobasidionannosum (Fr.) Bref s. str. были найдены только в одном случае на возвышенном месте на корнях елового вывала.
- Устойчивость сосны к корневым патогенам формируется еще в стадии жердняка, когда интенсивный рост сочетается с резкой дифференциацией и отпадом отстающих в росте деревьев. Деревья, успешно и без значительного снижения прироста прошедшие этот период, избежавшие «эффект группового угнетения», более устойчивы к корневой губке и опенку. Таким образом, при проведении лесовосстановительных работ в местах потенциального наличия инфекции корневых патогенов в почве следует стремиться к свободному произрастанию деревьев (обеспечивает высокую биологическую устойчивость сосны обыкновенной и препятствует распространению возбудителей гнили).
БИБЛИОГРАФИЧЕСКИЙ СПИСОК
- Ваганов, Е.А. Рост и структура годичных колец хвойных / Е.А. Ваганов, А.В. Шашкин. – Новосибирск: Наука, 2000. – 232 с.
- Емшанов, Д.Г. Методы пространственной экологии в изучении лесных экосистем. - Киев, Меркьюри Глоуб Юкрейн, 1999. -220 с.
- Изменение климата, 2007 г.: последствия, адаптация и уязвимость» Вклад Рабочей группы II в Четвертый доклад МГЭИК об оценках -106 с.
- Изменение климата, 2007 г.: физическая научная основа. Вклад Рабочей группы I в Четвертый доклад МГЭИК об оценках -163 с.
- Кузьмичев, В.В. Закономерности роста древостоев / В.В. Кузьмичев. – Новосибирск: Наука, 1977. – 135 с.
- Негруцкий, С.Ф. Корневая губка / С.Ф. Негруцкий. — М.: Агропромиздат, 1986, — 196 с.
- Негруцкий, С.Ф. Корневая губка / С.Ф. Негруцкий. — М.: Лесная промышленность, 1973, — 200 с.
- Обзор состояния и тенденций изменения климата России, Москва: Институт Глобального Климата и Экологии 2007. -23 с.
- Павлов, И.Н. Техногенные и биотические механизмы деструкции коренных лесов юга Сибири и их восстановление. автореферат диссертации на соискание ученой степени доктора биологических наук по спец: 06.03.03; 03.00.16. –М. 2007. -44 с.
- Павлов, И.Н. Закономерности образования очагов Heterobasidion annosum (Fr.) Bref. s. str. в географических культурах сосны обыкновенной (Минусинская котловина) / И.Н. Павлов [и др.] // Хвойные бореальной зоны. - 2008. – Т.25. – №. 1-2. – С. 28-36.
- Погребняк, П.С. Общее лесоводство / П.С. Погребняк. – М.: Колос, 1968. - 440 с
Почвенные факторы продуктивности сосняков (на примере Минусинских ленточных боров Красноярского края). – Новосибирск: Наука. – 1976. – 238 с.
- Почвы СССР. отв. ред. Г.В. Добровольский. - М. - Мысль, 1979. – 380 с.
- Романовский, М.Г. Продуктивность, устойчивость и биоразнообразие равнинных лесов европейской России / М.Г. Романовский. – М.: МГУЛ, 2002. – 97 с.
- Соколов, Д.В. Корневая гниль от опенка и борьба с ней / Д.В. Соколов. – М.: Лесная промышленность. – 1964. – 183 с.
- Стороженко, В.Г. Гнилевые фауты коренных лесов Русской равнины / В.Г. Стороженко. – М. – 2001. – 157 с.
- Уткин, А.И. Биологическая продуктивность лесов: Методы изучения и результаты / А.И. Уткин. - Лесоведение и лесоводство: Итоги науки и техники. – ВИНИТИ, 1975. –Т.1. – С. 9-139.
- Федоров, Н.И. Корневые гнили хвойных пород / Н.И. Федоров.– М.: Лесн. пром. – 1984. – 160 с.
- Шигапов З.Х. Генетический анализ природных популяций и лесосеменных плантаций сосны обыкновенной: автореф. дис. ... канд. биол. наук. 06.03.01. - Красноярск: Институт леса им. В.Н. Сукачева СО РАН, 1993. - 23 с.
- Burdon J.J. Epidemiology of damping of disease (Pythium irregulare) in relation to density of Lepidium sativum seedlings / J.J. Burdon, G.A. Chilvers // Annals of Applied Biology. – 1975. - № 81. – Р. 135-143.
- Connell J.H. Some mechanisms producing structure in natural communities: a model and evidence from field experiments / J.H. Connell // In: Ecology and Evolution of Communities M.L. Cody and J.M. Diamond eds., Belknap, Cambridge, Massachusetts. – 1975.
- Courtois, H. Influence of site factors on the germination of Fomes annosus / H. Courtois // European Journal of Forest Pathology. – 1972. - № 2. – Р. 152-171.
- Courtois, H. Influence of temperature, light and other factors on the mycelial growth of Fomes annosus / H. Courtois // Angewandte Botanik. – 1973. - № 47. – Р. 141-158.
- Cowling, E.B. Influence of temperature on growth of Fomes annosus isolates / E.B. Cowling, A. Kelman // Phytopathology. – 1964. - № 54. – Р. 373-378.
- Donaubauer, E. Die Bedeutung von Krankheitserregern beim gegenwartigen Eichensterben in Europa-eine Literaturubersicht / E. Donaubauer // Eur. J. Forest Pathol. - 1998. - 28, N 2. - P. 91-98.
- Guillaumin, J.J. Armillaria species in the northern temperate hemisphere / J.J. Guillaumin, С. Mohammed, S. Berthelay // In: Morrison, D.J., ed. Proceedings of the 7th international conference on root and butt rots; Vernon and Victoria, ВС. Victoria, ВС: International Union of Forestry Research Organization. August, 1988. – Victoria. – 1988. – Р. 27-43.
- Heterobasidion annosum: biology, ecology, impact and control /edited by S. Woodward et al. CAB International, Wallingford, UK. -1998. - 590 p.
- Hogg, E.H. Growth and dieback of aspen forests in northwestern Alberta, Canada, in relation to climate and insects / E.H. Hogg et all // Can. J. Forest Res 32. - 2002. - № 5. - P. 823-832.
- Hopper, R.J. Characterization of damage and biotic factors associated with the decline of Eucalyptus wandoo in southwest Western Australia / R.J. Hopper, K. Sivasithamparam // Can. J. Forest Res 35. - 2005. – N 11. - Р. 2589-2602.
- Gordon, J. Temperature and precipitation in Mongolia based on dendroclimatic investigations / J. Gordon et all // Pap. Workshop Southeast Asian DendrochronoL, Chiang Mai, 16-20 Febr., 1998. - IAWA Journal 20. - 1999. - №3. - P. 339-350.
- Korhonen, K. Fungi belonging to the genera Heterobasidion and Armillaria in Eurasia / K Korhonen // Грибные сообщества лесных экосистем: материалы координационных исследований – Москва-Петрозаводск, 2004. – Том 2. – С. 89-113.
- Korhonen, K. Interfertility and clonal size in the Armillaria mellea complex / К. Korhonen // Karstenia. – №18. – 1978. – P. 31-42.
- Nuorteva, P. The role of air pollution and climate change in development of forest insect outbreaks - guest editorial : Workshops "Eff. Possible Glob. Warming Insect Diversity and Distrib." and "Role Air and Soil Pollut. Dev. Forest Insect Outbreaks" during 20 Int. Congr. Entomol., Firenze, 25-31 Aug., 1996 / P. Nuorteva // Acta phytopathol. et entomol. hung 32. - 1997. - № 1-2. - P. 127-128.
- Redfern, D.B. Growth and behaviour of Armillaria mellea rhizomorphs in soil / D.B. Redfern // Transactions of the British Mycological Society. – 1973. – № 61. – Р. 569-581.
- Rigling, A. Waldfohrenbestande im Umbruch / A. Rigl-ing et all // Wald und Holz. - 1999. – 80. – № 13. - P. 8-12.
Rishbeth, J. The growth rate of Armillaria mellea / J. Rishbeth // Transactions of the British Mycological Society. – 1968. – № 51. – Р. 575-586.
- Rishbeth, J. Effects of soil temperature and atmosphere on growth of Armillaria rhizomorphs / J. Rishbeth // Transactions of the British Mycological Society. – 1978. – № 70. – Р. 213-220.
- Roll-Hansen, E. (1940) Undersokelser over Polyporus annosus Fr. saerlig med henblikk pa dens forekomst i Det sonnafjelske Norge / E. Roll-Hansen // Meddelelser fra det Norske Skogfor-soksvesen. – 1940. – NR 24. – № 7 (l). – Р. 1-100.
- Schwances, H.O., Courtois, H. and Ahrberg, H.E. () Okologie und Physiologie von Fomes annosus. In: Der Wurzelschwamm (Fomes annosus) und die Rotfaule der Fichte (Picea abies,). //Forstwissenschajiliche Forschungen, Beihefte zum Forstwissen-schaftliches Centralblatt, Verlag Paul Parey. - Hamburg und Berlin, 1976.- №36. PP. 14-30.
- Shaw III, C.G. Armillaria root disease. Agriculture handbook № 691 / C.G. Shaw III, G.A. Kile // Washington, USDA Forest Service – 1991.– 231 p.
- Technical Summary Working Group I to the Third Assessment Report of the IPCC // Geneva, Switzerland. – 2001. – P. 98.
- Thies, W.G. Laminated root rot in western North America / W.G. Thies, R.N. Sturrock // USDA Forest Service, General Technical Report PNW-GTR-349, 32 p.
- Vertui, F. Scots pine (Pinus sylvestris L.) die-back by unknown causes in the Aosta Valley, Italy / F. Ver-tui, F. Tagliaferro // Chemosphere 36. - 1998. - № 4-5. - P. 1061-1065.
- Woodward, S. Causes of decline in United Kingdom broadleaved stands / S. Woodward / Possible Limi-tation of Decline Phenomena in Broadleaved Stands. - Warsaw, 2006. - Р. 21-27.
|


