|
"Хвойные бореальной зоны" 2009г., №1, с. 48-57
Oсновная причина массового усыхания пихтово-кедровых лесов в горах Восточного Саяна – Корневые патогены
И.Н. Павлов, О.А. Барабанова, А.А. Агеев, А.С. Шкуренко, С.С. Кулаков, Д.В. Шпенглер, П.В. Губарев
ГОУ ВПО «Сибирский государственный технологический университет»
660049 Красноярск, пр. Мира, 82; e-mail: forester24@mail.ru
Установлена активизация процессов куртинного усыхания коренных темнохвойных лесов (Abies sibirica, Pinussibirica) в горах Восточного Саяна в последнее десятилетие. Гибель деревьев вызвана комплексом корневых патогенов (Heterobasidion parviporum Niemela & Korhonen, Armillaria mellea s.l., Phellinus sulphurascens Pilat.). Изменение климата, лесозаготовки, техногенное загрязнение – главные причины активизации патогенных свойств возбудителей корневых гнилей. Фактором, способствующим очаговому проявлению заболевания, является снижение биологической устойчивости древесных растений из-за ограничения роста корневых систем пихты и кедра на неглубоких почвах (корнеобитаемый слой 15-25 см), подстилаемых твердыми горными породами.
Ключевые слова: Heterobasidionparviporum Niemela & Korhonen; Armillariamellea s.l.; Phellinussulphurascens Pilat.; Восточный Саян
Drying of trees with increasing frequency has been noted last decade for native dark coniferous forests (Abies sibirica, Pinus sibirica) in the Eastern Sayan Mountains. Suggested that root rot fungal pathogens (Heterobasidion parviporum Niemela & Korhonen, Armillaria mellea s.l., Phellinus sulphurascens Pilat.) were the important cause of tree mortality. Climate change, woodcutting, technogenic pollution are the main reasons of the root rot pathogens activation. An additional important factor promoting the appearance of root rot disease is decreasing biological stability of trees because of limited growth of root systems of Siberian fir and Siberian pine in the insignificant root layer (15-25 centimeters) in conditions of spreading hard rocks under.
Key words: Heterobasidion parviporum Niemela & Korhonen; Armillaria mellea s.l.; Phellinus sulphurascens Pilat.; Eastern Sayan Mountains
Введение
Причины начавшегося в конце 20 века и возрастающего в настоящее время усыхания лесов (в первую очередь - хвойных) не установлены и часто объясняются комплексом почвенно-климатических факторов и антропогенным воздействием. К настоящему времени значительная деградация пихтово-еловых лесов отмечена на российском Дальнем Востоке (Манько, Гладкова, 1995, 2001; Манько и др., 1998; Власенко, 2005), пихтовых древостоев с примесью кедра, ели в районе озера Байкал (Мозолевская и др., 2003), горных пихтовых лесов в Кемеровской области (Алексеев, Шабунин, 2000). Затухая в одних местах, процесс дигрессии проявляется на других участках и в еще более значимых масштабах.
К началу 1990-х годов было выдвинуто более 170 рабочих гипотез, объясняющих усыхание леса (Rehfuess, 1991). И если до этого времени ведущая роль в ухудшении состояния древесных ценозов отводилась загрязняющим веществам, то по мере исследования на первое место, как и на начальном этапе изучения этого явления, ставятся погодные условия (Манько, Гладкова, 1995). При этом грибным инфекциям в массовом усыхании пихтово-еловых лесов не отводится главенствующей роли (только как сопутствующие факторы деградации) (Манько и др., 1998). В то же время авторы указывают на появление «в массе» плодовых тел опенка после рубок на пнях ели. Что, с нашей точки зрения, косвенно указывает на возможное патогенное воздействие именно Armillaria mellea s.l.
Причиной значительного занижения вредоносности возбудителей корневых гнилей является сложность их идентификации (часто отсутствие плодовых тел, обязательное сопряженное наличие следов поражения другими болезнями и вредителями). Так, при изучении воздействия двух следовавших друг за другом ураганов на старый лес с доминированием Fagus sylvatica и Abies alba в юго-восточной Словении роль корневых патогенов не рассматривается вообще (Nagel, Diaci, 2006). Хотя указанные симптомы - образование многочисленных просветов в пологе площадью до 1500 м2 в результате куртинной гибели деревьев после ураганов и выворот с корнями большинства деревьев - свидетельствуют о возможном развитии болезней корней.
ПРОГРАММА И МЕТОДИКА. ОБЪЕКТЫ
С конца ХХ века в горах Восточного Саяна наблюдается интенсификация процессов усыхания пихтово-кедровых лесов, значительно превышающая текущий естественный отпад. Гибель деревьев имеет ярко выраженный очаговый характер.
Очаги усыхания образуются при различной экспозиции склона. Куртинное усыхание не установлено на вершинах сопок, с очень мелким, быстро пересыхающим корнеобитаемым слоем (крайне неблагоприятные условия для развития корневых патогенов), а также на пониженных элементах рельефа с глубокими почвами (рис. 1).
Площадь очагов изменяется от 0,6 до 30 га (рис. 2). Усыханию подвержены деревья всех классов Крафта. Возраст погибших деревьев пихты – 70-160 лет (рис. 3). Возобновление темнохвойными породами в очагах - удовлетворительное. Гибель подроста пихты единична.
Очаговый характер поражения, развитие мицелия, характерные признаки деструкции ксилемы корней, основания ствола, истечение смолы позволили нам сделать предположение о воздействии корневых патогенов.
Исследования проведены на юго-западном склоне Восточного Саяна (Кизирское лесничество). Согласно Перечня лесорастительных зон и лесных районов Российской Федерации, утвержденного Приказом МПР России от 28.03.2007 г. № 68, территория относится к Южно-Сибирской горной зоне Алтае-Саянского горнотаёжного лесного района. Таксационная характеристика очагов усыхания представлена в таблице 1.
В естественных условиях обильное образование плодовых тел корневой губки встречается крайне редко (Федоров, 1984, Негруцкий, 1986), что во многом усложняет диагностику заболевания. Формирование плодового тела происходит при благоприятном стечении ряда факторов (увлажнение, затенение, температурный режим). Неблагоприятными условиями являются как жаркий, непродолжительный и зачастую засушливый летний период на территории сосняков Сибири, так и избыток влаги при недостатке тепла в темнохвойных лесах. Поэтому и плодовые тела гриба в темнохвойных лесах часто образуются на выворотах корней достаточно высоко над землей или даже, например, на торцевой части пня на высоте 1,5 м (рис. 4).
Для стимулирования образования плодовых тел корневой губки на исследуемой территории в начале вегетационного периода в очагах куртинного усыхания была произведена раскопка корней сильно ослабленных и усыхающих деревьев пихты и кедра. Раскопанные корни накрывались непрозрачным перфорированным полиэтиленом, ветвями, мхом, травой, древесным опадом. Таким образом, была смоделирована естественная среда образования плодовых тел корневой губки, образующаяся, например, при вывале деревьев или в норах землеройных животных. При этом следует иметь в виду, что избыток влаги так же вреден для образования базидиом как и её недостаток.
Для изучаемой территории характерны маломощные и среднемощные почвы. Наряду с описанием почвенных разрезов для изучения глубины корнеобитаемого слоя был изготовлен металлический щуп (по аналогии с «мечом Колесова»). Измерения проводились от центра очагов усыхания в нескольких радиальных направлениях с шагом 1-2 м.
ОБСУЖДЕНИЕ И АНАЛИЗ РЕЗУЛЬТАТОВ
При обследовании очагов куртинного усыхания найдено большое количество плодовых тел корневой губки (Heterobasidion parviporum Niemela & Korhonen), веера мицелия опенка (Armillaria mellea s.l.) под корой усыхающих деревьев и свежего сухостоя (рис. 5), а также плодовые тела малоизученного в России корневого патогена Phellinus sulphurascens Pilat. (рис. 6) с характерными признаками деструкции ксилемы Pinus sibirica Du Tour и Abies sibirica Ledeb. Четкой границы поражения древостоя разными корневыми патогенами не установлено.
P. sulphurascens Pilat. – возбудитель ламинатной корневой гнили (Laminated Root Rot), вызывающий расслоение по ранней древесине на слои толщиной в годичное кольцо, входящий в комплекс Phellinus weirii sensu lato. Впервые P. weirii (Murrill) Gilb. был обнаружен на Thuja plicata, на севере Айдахо (Murrill, 1914). В дальнейшем гриб также был обнаружен на Pseudotsuga menziesii, Abies spp., Picea spp. и Tsuga spp. (Bier and Buckland, 1947). В настоящее время по результатам скрещивания, а также молекулярных и серологических исследований принято существование двух интерстерильных групп P. weirii s.l.: - P. sulphurascens, поражающий Pseudotsuga menziesii и др. хвойные виды (Douglas-fir тип) и P. weirii (Murrill) Gilb. sensu stricto, развивающийся на Thuja plicata (red cedar тип) (Kotlaba, Pouzar 1970; Angwin, Hansen, 1993; Larsen, Cobb-Poulle, 1990; Larsen et al. 1994; Lim et al., 2005). Несмотря на выделение двух видов остается мнение, что использование названия P. weirii для всех грибов, вызывающих ламинатную гниль (в том числе и на Pseudotsuga menziesii) не может быть ошибочным (Thies, Sturrock, 1995).
Масштабы повреждения хвойных лесов P. weirii в Азии и западе Северной Америки чрезвычайно велики (Thies, Sturrock, 1995; Lim et al., 2008). В северной части штата Айдахо комплекс корневых болезней, включая ламинатную гниль, может покрывать сотни или даже тысячи гектаров лесов, образуя мозаично разреженный полог (Thies, Sturrock, 1995). Наибольший вред ламинантная гниль корней наносит североамериканским лесам из Pseudotsuga menziesii. Так, потери древесины в западных хвойных лесах США ежегодно составляют около 4,4 млн. м3 (Nelson et al., 1981). В штатах Вашингтон и Орегон 8 % коммерческих лесов подвержены заболеванию со снижением запаса древесины на 40-70 % (Goheen, Hansen, 1993)
В очагах усыхания за счет улучшения светового режима через разреженный полог отмечается стимулирование роста сеянцев и корневых отпрысков деревьев, так же, как и высокорослых кустарников, таких, как клен голый (Acer glabrum Torr.), холодискус разноцветный (Holodiscus discolor (Pursh.) Maxim.) и пузыреплодник (Physocarpus malvaceus) (Thies, Sturrock, 1995). Однако корни чувствительных сеянцев, растущих рядом с инфицированным деревом, могут быть поражены в течение нескольких лет.
По чувствительности древесные растения, произрастающие на западе Северной Америки, к воздействию возбудителей ламинатной гнили разделены на пять классов (Wallis, 1976; Filip, Schmitt, 1979; Hadfield, 1985; Nelson, Sturrock, 1993; Thies, Sturrock, 1995). К высокочувствительным отнесены:
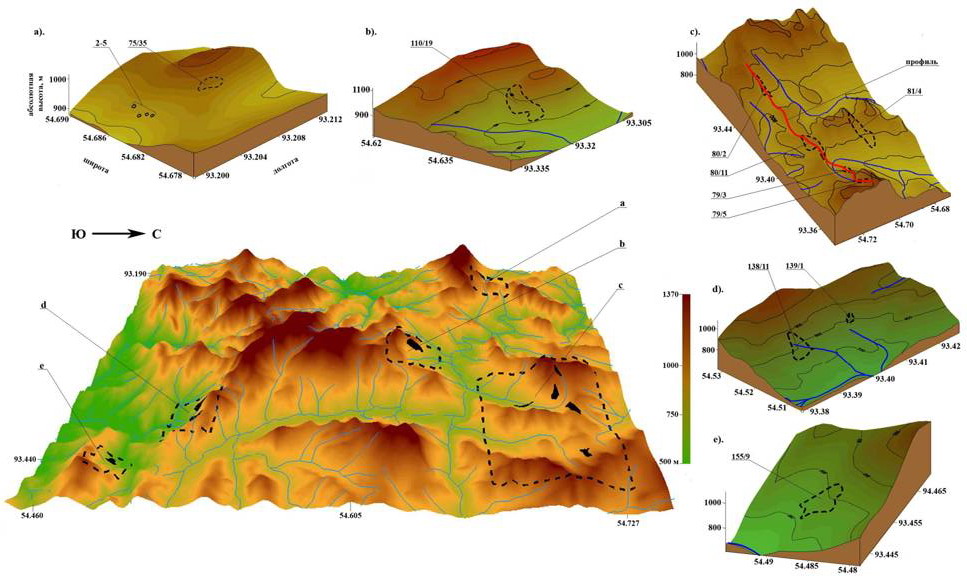 Рисунок 1 – Орографические закономерности образования крупных очагов куртинного усыхания Рисунок 1 – Орографические закономерности образования крупных очагов куртинного усыхания
|
|

|

|
Рисунок 2 – Очаг куртинного усыхания 75/35 |
Рисунок 3 – Погибший древостой Abies sibirica |

|

|
Рисунок 4 – Плодовое тело Heterobasidion parviporum Niemela & Korhonen на торцевой части пня на высоте 1,5 м |
Рисунок 5 – Веер мицелия Armillariamellea s.l. под корой усыхающего дерева Abies sibirica |

|

|
Рисунок 6 – Плодовое тело Phellinussulphurascens Pilat. |
Рисунок 8 – Прогалина среди пихтово-кедрового древостоя |
 Pseudotsugamenziesii (Mirb.) Franco; Abiesgrandis (Dougl. ex D. Don) Lindl.; Tsugamertensiana (Bong.) Carr.; Abies amabilis Dougl. ex Forbes; Abies concolor (Gord. & Glend.) Lindl. ex Hildebr. К среднечувствительным: Abies magnifica A. Murr.; Picea engelmannii Parry ex Engelm.; Sequoiadendron giganteum (Lindl.) Buchholz; Abies procera Rehd.; Taxus brevifolia Nutt.; Picea sitchensis (Bong.) Carr.; Abies lasiocarpa (Hook.) Nutt.; Tsuga heterophylla (Raf.) Sarg.; Larix occidentalis Nutt. К толерантным: Pinus contorta Dougl. ex Loud., Pinus lambertiana Dougl., Pinus monticola Dougl. ex D. Don. К стойким (resistant): Chamaecyparis nootkatensis (D. Don) Spach; Libocedrus decurrens Torr.; Pinus ponderosa Dougl. ex Laws.; Chamaecyparis lawsoniana (A. Murr.) Parl.; Sequoia sempervirens (D. Don) Endl.; Thuja plicata Donn ex D. Don. К невосприимчивым (immune): Acer macrophyllum Pursh.; Physocarpus malvaceus (Greene) Kuntze; Holodiscus discolor (Pursh) Maxim.; Alnus rubra Bong.; Acer glabrum Torr.; Acer circinatum Pursh. Pseudotsugamenziesii (Mirb.) Franco; Abiesgrandis (Dougl. ex D. Don) Lindl.; Tsugamertensiana (Bong.) Carr.; Abies amabilis Dougl. ex Forbes; Abies concolor (Gord. & Glend.) Lindl. ex Hildebr. К среднечувствительным: Abies magnifica A. Murr.; Picea engelmannii Parry ex Engelm.; Sequoiadendron giganteum (Lindl.) Buchholz; Abies procera Rehd.; Taxus brevifolia Nutt.; Picea sitchensis (Bong.) Carr.; Abies lasiocarpa (Hook.) Nutt.; Tsuga heterophylla (Raf.) Sarg.; Larix occidentalis Nutt. К толерантным: Pinus contorta Dougl. ex Loud., Pinus lambertiana Dougl., Pinus monticola Dougl. ex D. Don. К стойким (resistant): Chamaecyparis nootkatensis (D. Don) Spach; Libocedrus decurrens Torr.; Pinus ponderosa Dougl. ex Laws.; Chamaecyparis lawsoniana (A. Murr.) Parl.; Sequoia sempervirens (D. Don) Endl.; Thuja plicata Donn ex D. Don. К невосприимчивым (immune): Acer macrophyllum Pursh.; Physocarpus malvaceus (Greene) Kuntze; Holodiscus discolor (Pursh) Maxim.; Alnus rubra Bong.; Acer glabrum Torr.; Acer circinatum Pursh.
Несмотря на значительное число базидиоспор созревающих в базидиокарпах, их роль в образовании новых очагов инфекции невелика (Nelson, 1976). На инфицированных площадях патоген распространяется, главным образом, благодаря переходу мицелия от зараженных корней, контактирующих со здоровыми. По данным (Nelson, 1976), плодовые тела образуются достаточно редко. Однако нами в процессе обследования очагов куртинного усыхания пихтово-кедровых лесов в Восточном Саяне найдено большое количество плодовых тел P. sulphurascens, в основном - на выворотах корней пихты. После гибели инфицированных деревьев P. sulphurascens может существовать как сапрофит 50 и более лет на инфицированных пнях и больших корнях (Childs, 1963; Hansen, 1979). В прибрежных плантациях небольшие группы больных деревьев становятся заметны в 15-20-летнем возрасте, в то же время в континентальных районах очаги инфекции не проявляются ранее 20-30-летнего возраста (Thies, Sturrock, 1995).
При исследовании биоразнообразия грибов отмечено достаточно широкое распространение P. weirii sensu lato на территории бывшего СССР (ЭССР; Кижский архипелаг; Урал; Новосибирская, Томская, Иркутская области; Красноярский край, Дальний Восток) (Бондарцева, Пармасто, 1986; Бондарцева, Лосицкая, Руоколайнен, 1999). Плодовые тела были обнаружены на крупном валеже и пнях хвойных пород, на древесине и коре (Abies sibirica, Picea obovata, Larix sibirica, реже Pinus sibirica, P. silvestris). При этом патогенные свойства P. weirii s.l. исследованы не были.
При исследовании орографических закономерностей образования крупных очагов куртинного усыхания (инициированных корневыми патогенами) установлено их отсутствие на вершинах сопок (рис. 7). Исключение составляет участок 79/2, представляющий собой плато протяженностью около километра с понижением в центре. Высотным ограничением является отметка в 1 тыс. м над уровнем моря. Причина заключается в недостатке эффективной температуры для развития корневых патогенов. Только один кластер усыхания располагается на высоте 1000-1080 м. При этом недостаток тепла, определяемый высотой, компенсируется южной экспозицией очага. Температура является исключительно важным параметром, определяющим не только распространение корневых патогенов, но и их агрессивность. Так, рост ризоморф опенка происходит в диапазоне температуры почвы 5-28 0С (Rishbeth, 1968). Скорость распространения при средней температуре в верхнем слое (15 см) почвы 10 0С достигает 1,5 м в год. Даже незначительное повышение температуры приземного слоя воздуха в условиях недостатка тепла в горных условиях значительно влияет на патогенные свойства корневых патогенов.
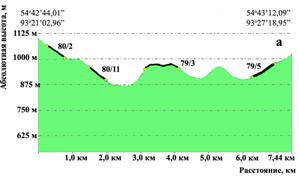
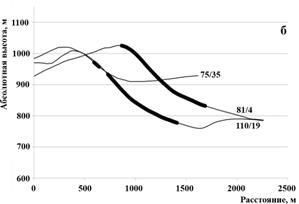
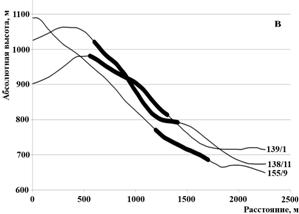
Рисунок 7 – Продольный профиль рельефа с нанесенными очагами куртинного усыхания (а- профиль, пересекающий очаги 80/2, 80/11, 79/3, 79/5, горизонтальная проекция показана на рис. 1с; б – очаги 75/35, 81/4, 110/19; в- очаги 138/11, 139/1, 155/9)
В пониженных элементах рельефа (в т.ч. в поймах ручьев) наблюдается единичная и групповая гибель деревьев пихты, ели, кедра (среди возбудителей корневых гнилей доминирует A. mellea s.l.).
Чаще всего очаги куртинного усыхания образуются в средней части хорошо освещенных склонов ЮВ-Ю-ЮЗ экспозиции.
Для исследуемой территории характерно наличие небольших прогалин площадью до 2 га. (рис. 8). При их обследовании установлена устойчивая закономерность – в местах отсутствия древесной растительности глубина возможного корнепроницаемого слоя, ограниченного выходом материнской породы, значительно меньше, чем под прилегающими древесными массивами (рис. 9).
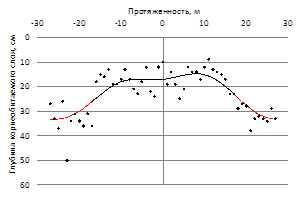
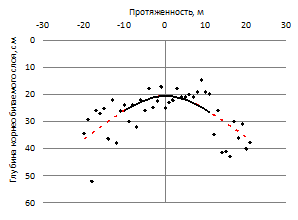
Рисунок 9 – Глубина корнепроницаемого слоя, ограниченного выходом материнской породы на прогалине естественного происхождения (направление профиля: а- запад-восток; б – север-юг)
При анализе закономерностей появления очагов усыхания на различных элементах рельефа Саян в сочетании с исследованием глубины корнеобитаемого слоя установлена устойчивая закономерность. На вершинах сопок, с очень мелким, быстро пересыхающим корнеобитаемым слоем (крайне неблагоприятные условия для развития корневых патогенов), а также на глубоких почвах (высокая устойчивость хозяина) очаги куртинного усыхания не образуются (рис. 10). Наиболее неблагоприятные для пихты и кедра условия складываются на неглубоких почвах (корнеобитаемый слой - 15-25 см), подстилаемых твердыми горными породами, не доступными для освоения корнями (рис. 11). В этих условиях снижение биологической устойчивости древесных растений из-за недостатка влаги и
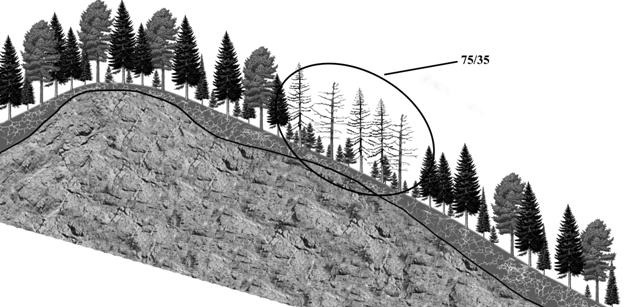
Рисунок 10 – Эдафические закономерности образования очага куртинного усыхания пихтово-кедрового древостоя (75/35)
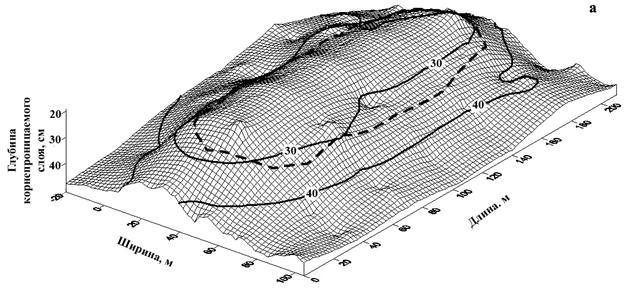
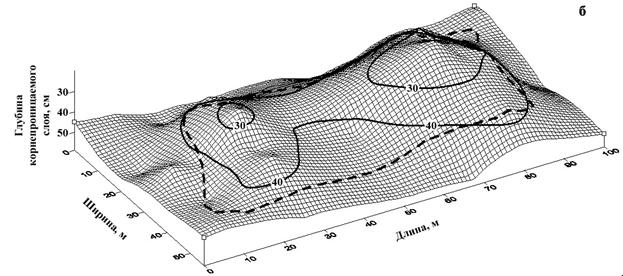
Рисунок 11 – Выход материнской твердой горной породы, ведущей к снижению глубины корнеобитаемого слоя: а – очаг 75/35; б – очаг 139/1
питательных веществ, хорошая прогреваемость почвы способствуют возрастанию вирулентности и агрессивности корневых патогенов. Как следствие – разрастание очага усыхания за пределы неблагоприятных эдафических условий.
Снижению биологической устойчивости пихтово-кедровых лесов также способствуют стволовые вредители в период их массового размножения (Monochamus urussovi Fisch.; Ips typographus L.; Pityogenes chalcographus L.). Возрастанию их численности способствует агрессивная лесозаготовительная деятельность в регионе.
ВЫВОДЫ
- Причиной усыхания пихтово-кедровых лесов является комплекс корневых патогенов (Heterobasidion parviporum Niemela & Korhonen, Armillaria mellea s.l., Phellinus sulphurascens Pilat.).
- Основными причинами активизации возбудителей корневых гнилей и последующего куртинного усыхания пихтово-кедровых лесов являются:
- увеличение количества и частоты осадков, рост приземной температуры воздуха;
- рубка деревьев, способствующая распространению корневых патогенов;
- рост ветровой нагрузки на леса (вследствие изменения климата и увеличения площадей вырубок), содействующий развитию раневых гнилей;
- рост численности популяций стволовых вредителей (Monochamus urussovi Fisch.; Ips typographus L.; Pityogenes chalcographus L.). Этому способствует потепление климата; лесозаготовки; деревья, ранее погибшие от возбудителей корневых гнилей. Значительно ослабленные корневыми патогенами деревья не могут обеспечить успешную защиту от насекомых и в период их массового размножения погибают.
- техногенное загрязнение, ведущее к общему снижению устойчивости хвойных лесов.
- Причиной, способствующей очаговому проявлению заболевания, является снижение биологической устойчивости древесных растений из-за ограничения роста корневых систем пихты и кедра на неглубоких почвах (корнеобитаемый слой 15-25 см), подстилаемых твердыми горными породами.
- В дальнейшем, при проведении восстановительных мероприятий, следует исключить посадку лесных культур на участках с неглубокими почвами, подстилаемыми твердыми горными породами. Для этого необходимо заранее, например, до проведения различных видов рубок, отметить места куртинного усыхания с помощью GPS. Очаги, в том числе и потенциальные, оставляются под естественное заращивание.
БИБЛИОГРАФИЧЕСКИЙ СПИСОК
- Алексеев, В.А. Побеговый рак пихты сибирской. Описание болезни и методические рекомендации по его полевой диагностике / В.А. Алексеев, Д.А. Шабунин. - СПб: СПб НИИЛХ, 2000. - 29 с.
- Бондарцева, М.А. Определитель грибов СССР. Афиллофоровые грибы / М.А. Бондарцева, Э.Х. Пармасто. -. – Л: Наука, 1986. - Вып. 1. - 180 с.
- Власенко, В.И. Усыхающие ельники среднего Сихотэ-Алиня / Власенко В.И. // Ритмы и катастрофы в растительном покрове российского Дальнего Востока: Материалы Международной научной конференции, Владивосток, 12-16 окт., 2004. - Владивосток, 2005. - С. 129-135.
- Манько, Ю.И. Мониторинг усыхания пихтово-еловых лесов в центральной Сихоте-Алине / Ю.И. Манько [и др.]. // Лесоведение. – 1998. - №1.– С. 3-15.
- Манько, Ю.И. О факторах усыхания пихтово-еловых лесов на Дальнем Востоке / Ю.И., Манько, Г.А. Гладкова // Лесоведение. – 1995. –№ 2. – С. 3-12.
- Манько, Ю.И. Усыхание ели в свете глобального ухудшения темнохвойных лесов / Ю.И., Манько, Г.А. Гладкова- Владивосток: Дальнаука, 2001. - 231 с.
- Мозолевская, Е.Г. Роль болезней и вредителей в ослаблении и усыхании пихты в Байкальском заповеднике в середине 80-х годов / Е.Г. Мозолевская, Т.В. Галасьева, Э.С. Соколова // Лесной вестник. – 2003. - №2. – С. 136-142.
- Негруцкий, С.Ф. Корневая губка / С.Ф. Негруцкий. — М.: Агропромиздат, 1986, — 196 с.
- Федоров, Н.И. Корневые гнили хвойных пород / Н.И. Федоров. – М.: Лесная промышленность, 1984. – 160 с.
Angwin, P.A. Pairing tests to determine mating compatibility in Phellinus weirii / P.A. Angwin, E.M. Hansen // Mycol Res 97, 1993.- P. 1469–1475.
- Bier, J.E. Relation of research in forest pathology to the management of second growth trees / J.E. Bier, D.C. Buckland / B.C. Lumberman, 1947.-№ 31(1). - Р. 49-51, 64, 66.
- Childs, T.W. Poria weirii root rot / T.W. Childs // Phytopathology, 1963. - №53. – Р. 1124–1127.
- Filip, G.M. Susceptibility of native conifers to laminated root rot east of the Cascade Range in Oregon and Washington / G.M. Filip, C.L. Schmitt // Forest Science. – 1979. - №25. – Р. - 261-265.
- Goheen, D.J. Effects of pathogens and bark beetles on forests / D.J. Goheen, E.M. Hansen // Beetle-pathogen interactions in conifer forests. - London: Academic Press. - 1993. - P. 175-196.
- Hadfield, J.S. Laminated root rot, a guide for reducing and preventing losses in Oregon and Washington forests / J.S. Hadfield.- Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Region. - 1985. -13 p.
- Hansen, EM Survival of Phellinus weirii in Douglas-fir stumps after logging / EM Hansen // Can J For Res. – 1979. – №9. – Р. 484–488.
- Kotlaba, F. Revision of the original material of Phellinus sulphurascens Pil., Xanthochrous glomeratus ssp. heinrichii Pil. and Polyporus rheades Pers. (Hymenochaetaceae) / F. Kotlaba, Z. Pouzar // Ceska Mykologie. – 1970. - №24. – P.146-152.
- Larsen, M.J. Phellinus (Hymenochaetaceae) / M.J. Larsen, L.A. Cobb-Poulle // A survey of the world taxa. Synop Fung. – 1990. - №3. – P. 1-206.
- Larsen, M.J. Phellinus sulphurascens and the closely related P. weirii in North America // M.J. Larsen, F.F. Lombard, J.W. Clark // Mycologia. – 1994. - №86. – P. 121-I30.
- Lim Y.W. Differentiating the two closely related species, P. weirii and P. sulphurascens / Y.W. Lim, Y.C.A. Yeung, R.N. Sturrock, I. Leal, C. Breuil // Forest Pathol. – 2005. - №35. – P. 305–314.
- Lim, Y.W. Distinguishing homokaryons and heterokaryons in Phellinus sulphurascens using pairing tests and ITS polymorphisms / Y.W. Lim, et. al. // Antonie Van Leeuwenhoek. – 2008. - №93. – P. 99-110.
- Murrill, W.A. An enemy of the western red cedar / Murrill, W.A. // Mycologia. – 1914. - №6. – P. 93-94.
- Nagel, T.A. Intermediate wind disturbance in an old-growth beech-fir forest in southeastern Slovenia / T.A. Nagel, J. Diaci // Can. J. Forest Res 36. - 2006. - № 3. - С. 629-638.
- Nelson, E.E. Colonization of wood disks initiated by basidiospores of Phellinus weirii (laminated root rot) / E.E. Nelson // For Sci. – 1976. - №22. – P. 407-411.
- Nelson, E.E. Laminated root rot of western conifers / E.E. Nelson, N.E. Martin, R.E. Williams // For. Insect Disease Leafl. 159. Washington, DC: U.S. Department of Agriculture. - 1981. - 6 p.
- Nelson, E.E. Susceptibility of western conifers to laminated root rot (Phellinus weirii) in Oregon and British Columbia field tests / E.E. Nelson, R.N. Sturrock // Western Journal of Applied Forestry. – 1993. - №8. – P. 67-70.
- Rehfuess, K.E. Review of forest decline research activities and results in the Federal Republic of Germany / K.E. Rehfuess // J. Environ. Sci. and Health. – 1991. - V 26. -№3. - P. 415-445.
- Rishbeth, J. The growth rate of Armillaria mellea / J. Rishbeth // Transactions of the British Mycological Society. – 1968. – № 51. – Р. 575-586.
- Thies, W.G. Laminated root rot in western North Americ / W.G. Thies, R.N. Sturrock // USDA Forest Service, General Technical Report PNW-GTR-349. - 32 p.
- Wallis, G.W. Phellinus (Poria) weirii root rot: detection and management proposals in Douglas-fir stands / G.W. Wallis // Forestry Tech. Rep. 12. Victoria, BC: Canadian Forestry Service, Pacific Forest Research Center. – 1976. -16 p.
|


