|
"Хвойные бореальной зоны" 2008г.,№3-4, с.
237-243
СОСТОЯНИЕ ПИХТОВО-КЕДРОВЫХ ЛЕСОВ ПРИРОДНОГО ПАРКА «ЕРГАКИ» И ИХ ФЛУОРЕСЦЕНТНАЯ ДИАГНОСТИКА
Третьякова И.Н.1, Бажина Е.В.1, Пахарькова Н.В.2, Сторожев В.Н.3
1Институт леса им. В.Н.Сукачева СО РАН
660036 Красноярск, Академгородок, 50; е-mail: genetics@ksc.krasn.ru
2ГОУ ВПО «Сибирский федеральный университет»
660041 Красноярск, пр. Свободный, 79
3Природный парк «Шушенский бор»,
660009 Красноярск, ул.Ленина, 123-а
Современными методами биоиндикации проводилась комплексная оценка пихтово-кедровых древостоев на территории природного парка «Ергаки» (Западный Саян). Выявлено, что сильно поврежденные биоценозы сосредоточены в районах среднегорья и высокогорья на склонах северной экспозиции. В данных биоценозах происходит интенсивное усыхание пихты сибирской: снижается длина хвои, появляются некрозы и хлорозы, падает урожайность шишек и качество семян, снижается фотосинтез. Данные ТИНУФ (термоиндуцированные изменения нулевого уровня флуоресценции) хлорофилла хвои показали, что пихта не имеет глубокого покоя в зимний период. Вероятно, этот феномен у пихты приводит к ее усыханию, вследствие потери влаги при транспирации.
Ключевые слова: парк «Ергаки», семенная продуктивность, качество семян, флуоресцентная диагностика
The value of fir-pine stands on territory of Natural Park “Ergaki” (Western Sayan) was conducted the modern methods of bioindication. It is revealed strong damage biocenoses were found in the middle and high mountains on the slopes of northern exposition. In these biocenoses goes the intensive drying of fir stands. In disturbance biocenoses the length of needle is decreased, necrosis and chlorosis are increased; the crop of female cones and seed quality and photosynthesis is decreased. TINUF date of fir and pine needles shown that fir-trees haven’t deep winter dormancy. It is possible that this phenomenon leads to its damage because moisture is losing at transpiration.
Key words: natural park “Ergaki”, bioindication, seed productivity, seed quality, fluorescentnja diagnostics
Введение
В настоящее время во многих регионах планеты происходит усыхание лесов (Johnson, Siccama, 1984; Forest decline., 1989, 1993). Это приводит к значительному уменьшению видового разнообразия, сокращению и исчезновению популяций и ценных форм лесных древесных видов. Предполагается, что в ближайшие десятилетия исчезнет около 25% ныне живущих видов. В числе видов, наиболее подверженных усыханию, оказались виды рода Abies(Mill.). Пихтовые леса интенсивно усыхают на территории ряда европейских стран, США, Канады, России, в Восточных Гималаях (Лесные экосистемы., 1990; Манько, Гладкова, 1995, 1999; Бажина, Третьякова, 2001; Landmann et.al., 1987; Forest decline., 1989, 1993). В горах Южной Сибири (Хамар-Дабан, Западный и Восточный Саян), начиная с семидесятых годов прошлого века, наблюдается интенсивное усыхание древостоев пихты сибирской (AbiessibiricaLedeb.) (Третьякова, Бажина, 1994, 1995).
Природный парк «Ергаки» - уникальный уголок природы в Западном Саяне, занимающий центральную часть федеральной трассы М54 Красноярск-Кызыл. Общая площадь природного парка "Ергаки" составляет 217 тыс. га. Парк создан, прежде всего,
___________________
*Работа поддержана РФФИ и ККФН (гранты 08-04-00107, 08-04-90001, 09-04-98000).
для охраны уникальных природных комплексов и объектов Западного Саяна, сохранения редких и исчезающих видов растений и животных, поддержания экологического баланса в условиях рекреационного использования территории. Основными лесообразователями в данном регионе являются пихта сибирская (Abiessibirica Ledeb.) и кедр сибирский (Pinussibirica Du Tour). В последние десятилетия на территории парка происходит интенсивное усыхание пихтовых лесов, начиная с высоты 700м над ур. м. и выше (Третьякова, Бажина, 1994, 1995; Третьякова и др., 1996). В связи с этим особенно актуально проведение мониторинга состояния лесных экосистем, выявление особенностей и возможных причин их неблагополучия. С этой целью выполнена комплексная оценка состояния пихтово-кедровых древостоев на территории природного парка «Ергаки» (Западный Саян) современными методами биоиндикации, в том числе флуоресцентной диагностикой.
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЙ
Исследования проводились в пихтово-кедровых древостоях Западного Саяна на 6 пробных площадях (п/п), заложенных в низко-, средне- и высокогорных районах вдоль Федеральной трассы № 54 Красноярск-Кызыл. Первая п/п расположена на высоте 450 м над ур.м. (район стационара Института леса, окрестности пос. Танзыбей), вторая п/п - в среднегорье (700-800 м над ур.м, северный макросклон хр. Кулумыс). Третья и четвертая п/п – на высоте 1450 м над ур.м. (северный макросклон хр. Ойский, оз. Ойское). Пятая - на высоте 1150 м (южный макросклон хр. Ергаки, турбаза Тушканчик), шестая – на южном макросклоне хр. Ергаки, 1514 м над ур.м. (оз. Светлое).
Определение содержания токсикантов (фтора, серы, тяжелых металлов) проводилось в хвое деревьев. Для этого на каждой п/п с 3 модельных деревьев кедра и пихты отбиралось 300 граммов хвои побегов 1-5 года жизни, хвоя высушивалась. Содержание токсикантов определялось в агрохимической лаборатории «Хакассия» (Методические указания, 1990, 1992, 1995, 2004).
На всех п/п было проведено описание жизненного состояния древостоев (Алексеев, 1989), измерены таксационные показатели деревьев и проведено описание морфоструктуры кроны модельных деревьев (Третьякова и др., 1996). Определялись основные морфометрические параметры побегов: линейный прирост побега, число хвоинок на 1см побега, длина хвои, процент хлорозов и некрозов хвои, а также семенная продуктивность шишек и качество их семян.
Для физиологической оценки состояния растений были использованы методы регистрации замедленной флуоресценции хлорофилла и термоиндуцированных изменений нулевого уровня флуоресценции хлорофилла (ТИНУФ), а также комплекс аппаратуры, разработанной на кафедре экотоксикологии и микробиологии Сибирского федерального университета (Гаевский и др., 1987). Регистрация замедленной флуоресценции (ЗФ) проводилась на компьютеризованном флуориметре «Фотон-10». В качестве показателя замедленной флуоресценции было взято отношение значений интенсивности быстрой и медленной компонент затухания свечения, измеряемых на высоком (120 вт/м2) и низком (10вт/м2) возбуждающем свету, соответственно (ОП ЗФ). Благодаря относительности измеряемой величины, данный показатель не зависит от массы и размера исследуемого растительного образца, а при подавлении фотосинтетической активности значительно снижается (Григорьев, Пахарькова, 2001).
ТИНУФ регистрировались на флуориметре «Фотон-5». Нагрев образцов, погруженных в воду, проводился в диапазоне от 20 до 80°С. В качестве показателя глубины покоя использовалось отношение интенсивностей быстрой флуоресценции Fо при 50°С и 70°С (коэффициент R) (Гаевский и др., 1987). Все данные обработаны статистически с помощью пакета анализа Microsoft Excel-2000.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Содержание фтора, серы и тяжелых металлов. Определение содержания в хвое кедра и пихты фтора, серы и тяжелых металлов в природном парке «Ергаки» показало, что его можно отнести к территории со слабым уровнем химического загрязнения. Тем не менее, в хвое кедра и пихты наблюдается накопление цинка и сульфатов. Содержание этих токсикантов приближалось к максимально допустимому уровню (МДУ) (табл. 1).
Таблица 1 – Содержание в хвое кедра и пихты фтора, серы и тяжелых металлов на территории природного парка «Ергаки»
Вид |
Содержание тяжелых металлов, мг/кг |
Фтор,
мг/кг |
Сульфаты
SO4,
мг/кг |
Zn |
Cu |
Cd |
Pb |
As |
Hg |
В пересчете на сухое вещество |
Пихта |
37,4±3,86 |
2,1±0,25 |
0,05±0,004 |
0,84±0,080 |
0,014±0,003 |
0,018±0,004 |
1,1±0,17 |
1050±300 |
Кедр |
44,0±16,6 |
1,8±0,09 |
0,11±0,018 |
0,49±0,022 |
0,01±0,002 |
0,01±0,001 |
2,0±0,33 |
1150±200 |
МДУ,
мг на кг натурального продукта |
50,0 |
30,0 |
0,3 |
5,0 |
0,5 |
0,05 |
20,0 |
1560 |
Жизненное состояние лесных биоценозов. Оценка жизненного состояния биоценозов в данном регионе показала, что оно различается в зависимости от условий произрастания (табл. 2). К классам биоценозов с начальной стадией повреждения относятся районы низкогорья (п/п № 1) и высокогорья (п/п №№ 5-6, расположенные на южном макросклоне хр. Ергаки). Сильно поврежденные биоценозы сосредоточены в районе среднегорья и высокогорья на макросклонах северной экспозиции (п/п №№ 2-4). Здесь происходит массовое усыхание деревьев пихты сибирской (Третьякова, Бажина, 1995).
Таблица 2 – Индекс жизненного состояния лесных биоценозов природного парка «Ергаки»
№
п/п |
Высота над ур. м. |
Состав древостоя |
Индекс жизненного состояния |
Класс лесных биоценозов |
Возраст,
м |
Диаметр на 1,3м, см |
1 |
450 |
6П4К |
81,1 |
Начальное повреждение |
60-120 |
9,8-16,5 |
2 |
800 |
6П4К |
36,4 |
Сильно поврежденный |
115-200 |
20,7-24 |
3 |
1450 |
8П2К |
66,6 |
Поврежденный |
115-200 |
4 |
1450 |
6П4К |
55,4 |
Поврежденный |
115-200 |
21,4-3,0 |
5 |
1130 |
7Е2К1П |
80 |
Начальное повреждение |
60-140 |
12,5-20 |
6 |
1514 |
6П4К |
81,5 |
Начальное повреждение |
100-200 |
17-24 |
Анализ морфоструктуры кроны и состояния генеративных органов модельных деревьев пихты сибирской, произрастающих в лесных экосистемах Западного Саяна, показал, что на п/п № 1, 5 и 6 деревья пихты в возрасте 60-120 лет, не имеют внешних признаков деградации. Протяженность генеративных ярусов у них занимает менее половины протяженности кроны. При этом женский генеративный ярус сосредоточен на вершине дерева и занимает 1,0-2,0 м. Ниже следует мужской генеративный ярус, протяженностью 1,5-4,0 м. На п/п №№ 2-4 возраст модельных деревьев составлял 115-145 лет. Протяженность женского генеративного яруса уменьшалась (0,55-2,0 м), а мужского– увеличилась (1,62-5,2 м). Деревья пихты здесь интенсивно усыхали - зона подверхушечного усыхания составляла 1,2-3,1м, что позволяет отнести их ко второй и третьей категории усыхания (Tretyakova, Bazhina, 2000).
Морфология побегов. Морфологические параметры побегов пихты и кедра, растущих в различных экологических условиях различается (табл. 3). В ненарушенных лесных биоценозах (п/п № 1) побеги деревьев, оказались несколько длиннее (на 5,7-23,4 %) и тоньше (на 20,7 %). Охвоенность побегов пихты сибирской стабильна, тогда как у кедра сибирского этот показатель в нарушенных сообществах высокогорья (п/п № 5, 6) увеличивается в два-три раза по сравнению с деревьями, произрастающими на п/п № 1. Неблагополучие экологической обстановки наглядно демонстрируют качественные признаки повреждения хвои (хлорозы и некрозы). В слабонарушенных сообществах отмечены точечные хлорозы, и некрозы, в нарушенных биоценозах площадь повреждения значительно увеличивается, у пихты преобладают некрозы пятнистого и мозаичного характера, у кедра - верхушечные хлорозы и некрозы.
Семенная продуктивность женских шишек и качество семян. Исследования семенной продуктивности макростробилов и качества семян пихты сибирской показали, что показатели структуры урожая женских шишек у пихты изменяются в зависимости от условий произрастания (табл. 4). Наиболее мелкие шишки формировались у деревьев, растущих в поврежденных биоценозах (п/п 3, 4). С понижением высоты местности в биоценозах с начальным повреждением показатели урожая семян женских шишек закономерно увеличиваются. Однако семенная продуктивность значительных изменений не претерпевала. Выход семян из шишек составлял 77-80 %.
Рентгенографический анализ семян пихты показал, что качество их в биоценозах с разной степенью повреждения крайне низкое (табл. 5). В поврежденных биоценозах высокогорья (п/п № 3-4) формировались только стерильные семена.
Таблица 3 – Характеристика вегетативных побегов и хвои
№ п/п |
Побеги |
Хвоя |
длина, |
диаметр, |
число хвои на |
длина,
мм |
ширина,
мм |
мм |
мм |
1см побега, шт |
пихта сибирская |
1 |
3,5±0,11 |
2,3±0,15 |
20±0,1 |
23,3±4,1 |
1,5±0,04 |
3, 4 |
3,3±0,11 |
2,9±0,12 |
19±0,2 |
19,7±0,49 |
1,6±0,20 |
кедр сибирский |
1 |
47,9±5,0 |
- |
5,9±1,20 |
48,0±11,0 |
- |
5 |
36,7±6,6 |
- |
12,1±1,70 |
52,0±0,60 |
- |
6 |
44,2±5,0 |
- |
20,0±0,5 |
60,0±0,80 |
- |
Таблица 4 – Семенная продуктивность макростробилов пихты сибирской
№
п/п |
Размеры шишек, мм |
Число семенных чешуй, шт. |
Число семян,
шт. |
Выход семян,
% |
длина |
ширина |
развитых |
общее |
развитых |
общее |
общее |
развитых |
1 |
53±5,7 |
22±0,6 |
103±25,2 |
129±28,8 |
171±32,8 |
199±38,6 |
77±2,3 |
66±2,2 |
3-4 |
37±0,9 |
10±0,3 |
115±3,0 |
138±3,2 |
201±7,2 |
220±7,5 |
80±2,0 |
73±1,9 |
Таблица 5 – Качество семян пихты сибирской
№ п/п |
Жизнеспособные |
Нежизнеспособные |
развитые |
недоразвитые |
всего |
поврежденные
конобионтами |
пустые |
всего |
1 |
1,91 |
35,7 |
54,8 |
11,1 |
34,2 |
45,2 |
3-4 |
0 |
0 |
0 |
0 |
100,0 |
100,0 |
Семенная продуктивность шишек кедра сибирского на всех пробных площадях достаточно высокая - 68,0 - 85,8 % (табл. 6). Семена формируются отличного качества, полнозернистость их достигает 100 %.
Таким образом, мониторинг состояния пихты сибирской в лесных биоценозах природного парка «Ергаки», показал, что в районах низкогорья (п/п № 1) и на южном макросклоне хр. Ергаки (п/п №№ 5-6) оно удовлетворительное. Сильно поврежденные биоценозы сосредоточены в районе среднегорья и высокогорья на склонах северной экспозиции (п/п № 3-4). Оценка, проведенная по признакам вегетативных и генеративных органов, показала, что усыхание пихты сибирской происходит по типовому сценарию: наблюдаются изменения в архитектуре кроны деревьев, появляется зона подверхушечного усыхания, увеличивается угол прикрепления ветвей, форма кроны из узкопирамидальной становится плакучей. Жизненное состояние кедра сибирского вполне удовлетворительное.
Фотосинтетическая активность. Содержание хлорофилла в хвое кедра и пихты в районе низкогорья выше, чем в высокогорье (рис. 1). Летом содержание пигментов в хвое кедра сибирского и пихты сибирской падает в нарушенных биоценозах (п/п 4). В октябре-ноябре различия в содержании хлорофиллов становятся менее заметными.
Таблица 6 – Семенная продуктивность макростробилов кедра сибирского
№ п/п |
Размеры шишек,
мм |
Число семенных чешуй, шт |
Число семян,
шт |
Выход семян,
% |
длина |
ширина |
развитых |
общее |
развитых |
общее |
общее |
развитых |
1 |
67±0,9 |
45±0,9 |
52±0,9 |
70±1,5 |
88±1,7 |
- |
- |
85,8±2,58 |
3-4 |
55±0,8 |
45±0,7 |
53±0,7 |
70±0,9 |
91±0,9 |
96±1,02 |
68±0,7 |
31±0,6 |
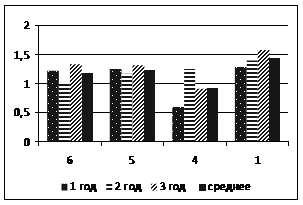 
а
б
Рисунок 1 – Содержание хлорофиллов на различных п/п (№№ 1,4-6) в одно- , двух-, трехлетней хвое пихты сибирской (а) и кедра сибирского (б). Слева - в сентябре, справа - в январе
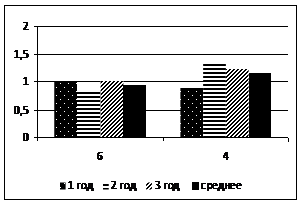
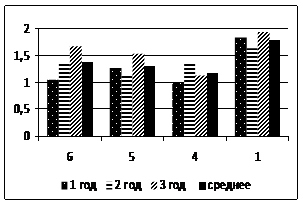
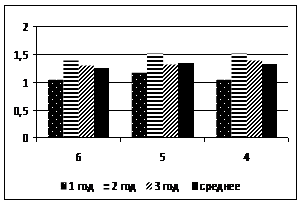
Относительный показатель замедленной флуоресценции (ОП ЗФ) в летнее время (июль) в высокогорье (п/п № 5 и 6) у кедра и пихты выше, чем в низкогорье (рис. 2). При этом ОП ЗФ у пихты снижается с увеличением высоты над уровнем моря. В поврежденном биоценозе (п/п № 4) этот показатель не превышал 4,0 (рис. 3).
Начиная с октября, интенсивность фотосинтеза, регистрируемая по параметрам замедленной флуоресценции, падает, причем быстрее в хвое, собранной на п/п № 4 и 6, расположенных на большей абсолютной высоте. Возможно, это связано с более коротким вегетационным периодом в горах, когда деревьям необходимо быстрее подготовиться к последующему периоду зимнего покоя. При положительных и невысоких температурах в эксперименте в хвое подавляется переменная флуоресценция хлорофилла и изменяется температурная зависимость выхода флуоресценции на уровне Fо: максимум около +50°С подавляется, и появляется новый максимум около +70°С (Гаевский, авторское свидетельство от 15.08.87). Коэффициент R является отношением значений низкотемпературного и высокотемпературного максимумов. При R меньше 1
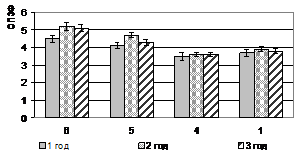
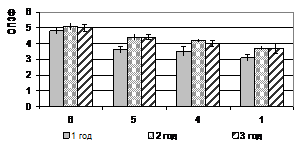
Рисунок 2 – Относительный показатель замедленной флуоресценции (ОПЗФ) хвои кедра сибирского (а) и пихты сибирской (б) на п/п №№ 1, 4-6, (июль, 2007)
растения находятся в состоянии покоя, при R больше единицы – фотосинтетически активны. Значения ОП ЗФ кедра сибирского и пихты сибирской в поврежденных биоценозах (п/п № 4) в декабре низкие, но глубокий покой у этих видов не наступает. При этом ОП ЗФ у пихты сибирской в нарушенных биоценозах выше, чем у кедра.
При искусственном выведении побегов хвойных из состояния зимнего покоя в лабораторных условиях в ноябре-декабре (рис. 4) можно отметить, что пихта сибирская значительно быстрее восстанавливает фотосинтетическую активность. Этот феномен свидетельствует о том, что пихта не имеет глубокого покоя в зимний период. Не исключено, что отсутствие покоя у пихты может способствовать ее усыханию в зимний период при кратковременных оттепелях и меньшей устойчивости к неблагоприятным факторам среды, например, к загрязнению воздуха, по сравнению с кедром сибирским. Определение ТИНУФ зимой в более поздние сроки (январь) показало, что в период резкого понижения температуры в горах Западного Саяна глубина покоя кедра сибирского и пихты сибирской становится одинаковой; выход из состояния покоя в лабораторных условиях происходит на шестые сутки.
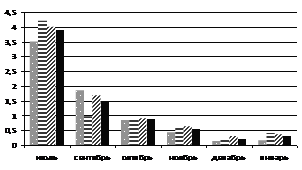
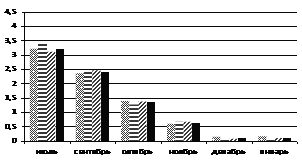
 Рисунок 3 – Относительный показатель замедленной флуоресценции хвои пихты сибирской (а) и кедра сибирского (б)
Рисунок 3 – Относительный показатель замедленной флуоресценции хвои пихты сибирской (а) и кедра сибирского (б)
Исследования состояния лесных фитоценозов на территории природного парка «Ергаки» показали, что леса данного экорегиона в горах следует отнести к поврежденным лесным экосистемам. Ранее показано, что одним из факторов, оказывающих негативное влияние на горные леса Южной Сибири, является загрязнение атмосферы (Третьякова и др., 1996). Однако, по данным химического анализа образцов хвои пихты и кедра, содержание токсикантов у них не превышает допустимых значений. Наблюдается лишь некоторое накопление цинка и сульфатов.
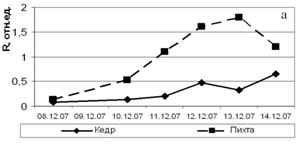
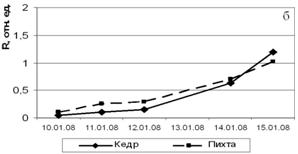
Рисунок 4 – Динамика ТИНУФ хвои кедра сибирского и пихты сибирской при выведении побегов из состояния покоя в лабораторных условиях при температуре 22°С и освещении лампами дневного света в декабре, 2007г (а) и в январе 2008г (б)
Другим не менее мощным фактором, оказывающим влияние на живые организмы, является глобальное потепление климата, происходящее и на территории Сибири (Tchebakova, Parfenova, 2003). Показано, что изменение климата оказывает влияние на ход роста и отпад деревьев, взаимоотношение видов, динамику древостоев и их возобновление (Павлов, Миронов, 2003; Tchebakova, Parfenova, 2003). Выявлено, что в результате погодно-климатических изменений, главным образом, за счет увеличения вегетационного периода в осеннее время произошел сдвиг в развитии генеративных органов у лиственницы сибирской и сосны обыкновенной (Романова, Третьякова, 2003; Третьякова, Носкова, 2004). У лиственницы сибирской в течение осени-зимы прошло завершение мейоза при микроспорогенезе и даже наблюдалось высыпание пыльцы при оттепелях зимой. У сосны обыкновенной микроспорогенез начинался не весной, а осенью, что не характерно для данного вида. Смещение сроков в развитии генеративных органов у представителей хвойных приводит к слабым урожаям женских шишек и семян и, даже, к полному их отсутствию.
Снижение репродуктивного потенциала и падение качества семян у пихты сибирской, возможно, свидетельствует о влиянии климатических изменений и на данный вид. Исследования показали, что ОП ЗФ кедра сибирского и пихты сибирской в поврежденных биоценозах в декабре низкие, но глубокий покой у этих видов не наступает. При этом у пихты сибирской этот показатель выше, чем у кедра сибирского. Флуоресценция хвои у пихты сибирской наблюдается даже в декабре.
Данные ТИНУФ хвои пихты и кедра также свидетельствуют о том, что пихта сибирская не имеет глубокого зимнего покоя. Данный вид находится в состоянии вынужденного покоя. Ранее было показано, что в условиях техногенного загрязнения сосна обыкновенная переходит к состоянию зимнего покоя в более поздние сроки и раньше выходит из него по сравнению с фоновыми древостоями (Григорьев, Пахарькова, 2001). При этом глубина покоя на протяжении всего зимнего периода значительно меньше. Известно, что при переходе растений к состоянию зимнего покоя в их тканях наблюдается ряд важнейших изменений, обеспечивающих их выживание в жестких условиях зимой (Горышина, 1989; Гаевский и др., 1991; Ходасевич и др., 1980; Шавнин, Фомин, 1993). По мнению Ю.С. Григорьева и Н.В. Пахарьковой (2001), неполный уход в покой деревьев из районов с высоким уровнем техногенного загрязнения приводит к водному дефициту и их сильному повреждению в зимний период.
Как показали исследования, на состояние биоценозов большое влияние оказывают экспозиционные различия склонов. Установлено, что в горах Южной Сибири различия северных и южных склонов по тепло- и влагообеспеченности могут быть настолько велики, что могут привести к резкому изменению типов растительности (в умеренно влажных районах), либо продуктивности и групп типов леса (во влажных районах) (Поликарпов и др., 1986). На южных склонах большая часть радиационного баланса расходуется на нагревание подстилающей поверхности, тогда как северные склоны и горизонтальная поверхность прогреваются меньше, а основная часть тепла расходуется на испарение. Вероятно, в этих условиях отсутствие зимнего покоя у пихты сибирской, вызванное потеплением климата, приводят к ее усыханию вследствие потери влаги при транспирации и преждевременному выходу из состояния покоя при кратковременных оттепелях.
БИБЛИОГРАФИЧЕСКИЙ СПИСОК
Алексеев, В.А. Диагностика жизненного состояния деревьев и древостоев/ В.А. Алексеев // Лесоведение. - 1989. - № 4. - С. 51–57.
Бажина, Е.В. К проблеме усыхания пихтовых лесов / Е.В. Бажина, И.Н.Третьякова// Успехи современной биологии. – 2001. - Т.121. - № 6. - С. 626-631.
Гаевский, Н.А. Сезонные изменения фотосинтетического аппарата древесных и кустарниковых растений / Н.А. Гаевский, Г.А. Сорокина, В.М. Гольд, И.В. Миролюбская// Физиология растений. - 1991. - Т. 38. - Вып. 4. - С.685-691.
Гаевский, Н.А. Способ определения степени глубины покоя древесных растений / Н.А. Гаевский [и др.]. - Авторское свидетельство №1358843 от 15.08.87.
Горышина, Т.К. Фотосинтетический аппарат растений и условия среды / Т.К. Горышина. - Л.:ЛГУ, 1989. - 204с.
Григорьев, Ю.С. Влияние техногенного загрязнения воздушной среды на состояние зимнего покоя сосны обыкновенной / Ю.С. Григорьев, Н.В. Пахарькова // Экология .- 2001.- № 6. С. 471-473.
Лесные экосистемы и атмосферное загрязнение /(под ред. В.А. Алексеева). - Л.: Наука, Ленинградское отделение, 1990. - 200с.
Манько, Ю.И. О факторах усыхания пихтово-еловых лесов на Дальнем Востоке / Ю.И. Манько, Г.А. Гладкова // Лесоведение. - 1995. - № 2. - С.3-12.
Манько, Ю.И. Об усыхании темнохвойных лесов в Северной Америке / Ю.И. Манько, Г.А. Гладкова // Лесоведение. - 1999. - № 5. - С. 56-62.
Методические указания по ионометрическому определению фтора в растительной продукции, кормах и комбикормах [Текст] - М: ЦИНАО (ВНИИА). - 1995г.
Методические указания по определению ртути в пробах пищевого сырья и продуктов (МЦК №5178-90) [Текст]. - 1990.
Методические указания по определению серы в растениях и кормах растительного происхождения [Текст] - М: ФГНУ «Росинформагротех».- 2004 г.
Методические указания по определению тяжелых металлов (цинк, медь, свинец, кадмий) в почвах и продукции растениеводства [Текст].- М.- 1992 г.
Павлов, И.Н. Динамика посевных качеств семян Larix sibirica Ledeb. в насаждениях юга Сибири / И.Н. Павлов, А.Г. Миронов // Хвойные бореальной зоны. - 2003. - Вып. 1. - С. 14-21.
Поликарпов, Н.П. Климат и горные леса Южной Сибири / Н.П. Поликарпов, Н.М. Чебакова, Д.И. Назимова. - Новосибирск: Наука, 1986. - 226с.
Романова, Л. И. Перспектива использования лиственницы в озеленении городов Сибири / Л. И. Романова, И. Н. Третьякова // Хвойные бореальной зоны. - 2003. - Вып. 1 (Лиственница). - С. 40-46.
Третьякова, И.Н. Влияние загрязнения среды окислами серы на морфоструктуру кроны, генеративную сферу и жизнеспособность пыльцы у пихты сибирской в Байкальском регионе / И.Н. Третьякова, О.Н. Зубарева, Е.В. Бажина // Экология. - 1996. - № 1. - С.17-23.
Третьякова, И.Н. Жизнеспособность пыльцы пихты сибирской в нарушенных лесных экосистемах гор Южной Сибири / И.Н. Третьякова, Е.В. Бажина // Экология.- 1994. - № 6. - С. 20-28.
Третьякова, И.Н. Морфоструктура кроны и состояние генеративной сферы у пихты сибирской в нарушенных лесных экосистемах близ озера Байкал / И.Н. Третьякова, Е.В. Бажина // Известия РАН. Сер. биол. - 1995. - № 6. - С. 685-692.
Третьякова, И.Н. Пыльца сосны обыкновенной в условиях экологического стресса / И.Н. Третьякова, Н.Е. Носкова // Экология. - 2004. - № 1. - С. 26-33.
Ходасевич, Э.В. Особенности регенерации фондов хлорофиллов a и b у желтеющих хвойных / Э.В. Ходасевич, А.И. Арнаутова, В.Н. Гвардиян // Физиология растений. - 1980. - Т. 27. - Вып. 4. - С.685-692.
Шавнин, С.А., Фомин Ф.С. Сезонные изменения флуоресценции хлорофилла хвои сосны обыкновенной / С.А. Шавнин, Ф.С. Фомин // Физиология растений. - 1993. - Т. 40. - № 2. - С.209-213.
Bonneau, M. Le deperissement du sapin pectine et de l'epicea commun dans le massif forestier vosgien est-il en relation avec l'etat nutritionnel des peuplements?/ G., Bonneau, M. Adrian // Rev. forest. fr. - 1987. - V. 39. - P. 5-11.
Forest decline and air pollution: a study of Spruce (Picea, Abies) on acid soils/ Ed. Schulze D.E.B. - Berlin. Heidelberg. New York. London. Paris: Springer-Verlag, 1989. - 475p.
Forest Decline in the Atlantic and Pacific Region/ R.F. Huettl, D. Mueller-Dombois (eds.). - Berlin, Heidelberg: Springer-Verlag, 1993. - 366 p.
Johnson, A.H. Decline of red spruce in the Northern Appalachians: assesing the possible role of air deposition / A.H. Johnson, T.G. Siccama //TAPPY journal. - 1984. - V. 67. - N 1. - P. 63-72.
Tchebakova, N. M. Stand growth and productivity of mountain forests in Southern Siberia in a Changing Climate / N. M. Tchebakova, E. I. Parfenova // CAB International. – 2003. – P. 189-197.
Tretyakova, I.N., Bazhina E.V. Structure of crown as well as pollen and seed viability of fir (Abies sibirica Ledeb.) in disturbed forest ecosystems of the Khamar-Daban Mts / I.N. Tretyakova, E.V. Bazhina // Ecology (Bratislava). – 2000. - V 19. - N 3. - Р. 280-294.
Поступила в редакцию 19 марта 2008 г.
Принята к печати 27 августа 2008 г. |


