|
"Хвойные бореальной зоны" 2007г.,№4-5, с. 487-491
Сезонные изменения содержания воды в меристематических тканях почек Picea obovata L. и Pinus sylvestris L. и её распределение в клетках
Алаудинова Е.В., Симкина С.Ю., Миронов П.В.
ГОУ ВПО «Сибирский государственный технологический университет»
660049 Красноярск, пр. Мира, 82; е-mail: simkina_svetlana@mail.ru; nvn@sibstu.kts.ru
Приведены данные о характере процессов обезвоживания и набухания меристематических тканей почек Picea obovata L. и Pinus sуlvestris L., протекающих в условиях естественного сезонного изменения температур. Установлено, что процесс снижения содержания воды в клетках меристем начинается значительно раньше осеннего снижения температуры воздуха и достигает в меристемах обеих пород в состоянии низкотемпературной устойчивости приблизительно одинакового минимального уровня (1,3?1,4 г воды/г а.с.м. ткани). Кроме того, вплоть до минус 40 °С в клетках меристематических тканей в условиях внеклеточного и внеорганного льдообразования сохраняется жидкая фаза воды, что, по-видимому, объясняет возможность протекания в этих условиях разнообразных биохимических процессов.
Adduce the results datas of study dehydrating and swellings meristematic tissues of vegetative buds of Picea obovata L. and of Pinus sуlvestris L. in the processes in conditions of natural seasonal change of temperatures. It is es-tablished that reduction of water contents in cells of meristema tissue stars long time before air temperature reduction in autumn. In low temperatured stability condition in both spectes of wood water content reaches approximately equal minimal rate (1,3?1,4 gram of water per gram of c.d.m. tissue). Becides right up to minus 40 °С in cases of extracellu-lar and extraorganic ice forming a the liquid phase of water keeps on. Apparently this fact explain an opportunity of Processing various biochemical processes in such conditions.
В последние годы всё более осознается роль северных лесов в сохранении и оздоровлении окружающей среды. В условиях бореальной зоны преобладающим элементом лесных биоценозов являются хвойные древесные растения.
Однако в условиях Восточной Сибири, характеризующихся резкой сменой времён года, благоприятный для роста период короток. При этом основные гистогенетические центры хвойных, в том числе и меристемы фотосинтетического аппарата, большую часть года остаются неактивными. Адаптируясь к сезонному снижению температуры окружающей среды, растения, относящиеся к эктотермным организмам, для сохранения своих живых клеток и возобновления вегетации в следующем году, реализуют различные механизмы низкотемпературной устойчивости. Несмотря на большие успехи, достигнутые в познании феноменологии и механизмов холодовой адаптации растений, необходимо отметить, что основное внимание исследователей сфокусировано сегодня на их биохимических и молекулярно-генетических аспектах. Однако необходимо отметить, что в проблеме криорезистентности и сохранения жизнеспособности при низких температурах большую роль играет поведение воды - уникального компонента биосистем, одновременно являющегося средой и непосредственным участником метаболических процессов. Очевидно, что биохимические изменения к клетках при адаптации к низким температурам в первую очередь направлены на изменение состояния воды, при котором её фазовые переходы становятся относительно безопасными.
Известно, что многолетние древесные растения, произрастающие в районах холодного и умеренного климата, адаптируясь к снижению температуры, испытывают сильное обезвоживание, предотвращая тем самым всегда летальное последствие фазового перехода вода-лёд при внутриклеточной кристаллизации [1-3].
На сегодняшний день среди исследователей нет окончательного мнения о безопасном уровне содержания воды в живых растительных клетках при отрицательных температурах окружающей среды. Имеющиеся данные варьируют в очень широких пределах. Кроме того, отсутствуют динамические характеристики процесса обезвоживания, протекающего в условиях естественного сезонного снижения температур, и количественного распределения оставшейся влаги внутри живых клеток.
В связи с этим в данной работе была поставлена задача изучить изменения в содержании и распределении воды в меристемах почек представителей семейства сосновых (Pinaceae) ели сибирской (Picea obovata L.) и сосны обыкновенной (Pinus sуlvestris L.), относящихся к наиболее морозостойким видам древесных растений, в начальный период холодовой адаптации осенью в условиях низких температур и возможных фазовых переходов воды зимой и весной вплоть до распускания почек. Изучение таких изменений важно для понимания особенностей низкотемпературной адаптации осенью.
Объектом исследования послужили деревья сосны обыкновенной и ели сибирской 55-60 летнего возраста, произрастающие в пригородной зоне города Красноярска в естественных древостоях на серых лесных и дерново-подзолистых почвах. Климат Красноярска и его окрестностей резко континентальный. Большие колебания температур, относительно малое количество осадков и низкая влажность воздуха весной и летом являются характерными особенностями красноярского климата. В городе и его окрестностях последний весенний заморозок обычно наблюдается в конце апреля. Осенью заморозки начинаются в третьей декаде августа, а в иные годы только в начале октября. Зимой устанавливается малооблачная сухая с сильными морозами погода [4].
Выбор в качестве объекта исследования меристематических тканей (зачаточных тканей побега и хвои в почках) не случаен, т.к., обладая большой чувствительностью к условиям перезимовки, именно они, в конечном итоге определяют выживание дерева в целом. Кроме того, при выделении меристем из почек удаётся получить весьма однородные по содержанию однотипных живых клеток образцы, что делает интерпретацию результатов более надёжной.
Отбор меристематических тканей проводили следующим образом: меристемы вегетативных почек срезали скальпелем по границе с ксилемой после удаления коры и почечных чешуй с побегов последнего года.
Для получения препаратов клеточных мембран и водорастворимых веществ клеток меристемы гомогенизировали с добавлением охлажденной до 0 °С дистиллированной воды в соотношении 1:20. Для охлаждения клеточных стенок гомогенат в течение 3 мин центрифугировали при 500 g. Супернатант, содержащий клеточные органеллы, их фрагменты и клеточные мембраны, центрифугировали в течение 30 мин при 22 000 g. Полученную модельную систему в дальнейшем называли комплексом клеточных мембран, а надосадочную жидкость – водорастворимыми веществами [5].
Методом гель-хроматографии супернатант, содержащий водорастворимые вещества клеток, разделяли на высокомолекулярную (белки) и низкомолекулярную фракции [6].
Содержание воды в тканях меристем определяли весовым методом. Для этого собранные побеги предварительно выдерживали в течение суток при температуре плюс 2 °С для регидратации тканей.
Сушку образцов проводили в вакуумном су-шильном шкафу при 80 °С в присутствии пятиокиси фосфора.
После длительного выдерживания (15 суток) в герметизированных бюксах определяли значение равновесного со льдом водосодержания для образцов растворимых веществ, комплекса клеточных мембран и меристематических тканей при различных отрицательных температурах.
Для предотвращения кристаллизации воды образцы массой 30-50 мг предварительно подсушивали в лиофилизаторе без замораживания до 30%-й влажности. За окончательный результат принимали среднеарифметическое значение водосодержания четырёх образцов, при этом погрешность составляла не более 3% [7].
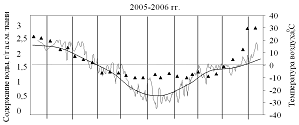
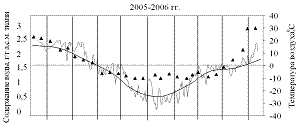
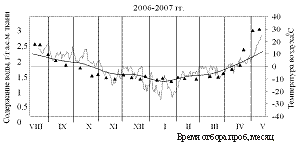
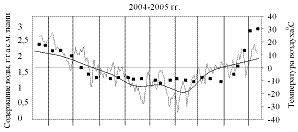
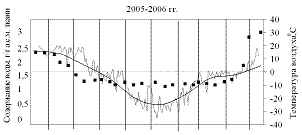
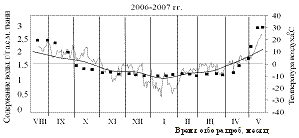
На рисунках 1 и 2 приведены результаты трехлетних наблюдений за сезонными изменениями влажности меристем почек ели и сосны и изменениями среднесуточных и среднемесячных температур воздуха сезонов 2004-2005, 2005-2006 и 2006-2007 гг.
Анализ результатов показал, что в меристемах почек обеих пород в августе при положительных среднесуточных температурах содержание воды составляло около 2,55±0,13 г/г а.с.м. ткани. Снижение влажности в тканях ели и сосны начиналось одновременно в середине сентября еще при положительных температурах воздуха.
В октябре (начальный период действия неблагоприятных температур), несмотря на то, что среднемесячные температуры воздуха еще оставались положительными, по утрам уже отмечались заморозки. Водосодержание меристем почек обеих пород продолжало снижаться, однако темпы снижения у сосны были несколько выше и поэтому минимальное среднезимнее содержание воды, составлявшее не более 1,31±0,05 г/г а.с.м. ткани, в меристемах почек этой породы устанавливалось в первой декаде ноября, что на две недели раньше, чем у ели.
Температурные условия в течение трёх лет наблюдений различались, хотя были довольно типичными.
^ содержание воды в меристемах почек ели
 изменения среднемесячных температур воздуха изменения среднемесячных температур воздуха
 изменения среднесуточных температур воздуха изменения среднесуточных температур воздуха
Рисунок 1 – Сезонные изменения влажности меристем почек ели
В Красноярске и его окрестностях август 2006 года (фаза формирования почек) характеризовался
¦ содержание воды в меристемах почек сосны
 изменения среднемесячных температур воздуха изменения среднемесячных температур воздуха
 изменения среднесуточных температур воздуха изменения среднесуточных температур воздуха
Рисунок 2 – Сезонные изменения влажности меристем почек сосны
более низкими среднемесячными и абсолютными температурами воздуха по сравнению с этими месяцами 2004 и 2005 гг.
Сентябрь также оказался самым холодным в 2006 году. Зима 2006-2007 гг., напротив, отличалась мягкостью, среднемесячные температуры составляли минус 12±2 °С. В 2004 году, август и сентябрь были тёплыми, но в октябре, хотя среднемесячные температуры оставались всё ещё положительными, уже наблюдалось понижение среднесуточных температур до минус 10 °С. Кроме того, самым холодным месяцем зимы 2004-2005 гг. был февраль.
Зима 2005-2006 гг. была наиболее суровой, среднемесячные температуры составляли ниже минус 20 °С, а среднесуточные иногда опускались до минус 35 °С.
Самая теплая и ранняя весна, согласно данным среднемесячной температуры воздуха, была в 2005 году, однако именно здесь зафиксированы максимальные и минимальные абсолютные температуры. Самая поздняя весна была в 2006 году.
Несмотря на определённые отличия температурных условий исследуемого периода (осень-зима-весна), зафиксированные в ходе трёхлетних наблюдений, уровень обезвоживания меристематических тканей почек обеих пород в состоянии низкотемпературной устойчивости достигал приблизительно одинаковых значений: содержание воды в клетках меристем осенью снижалось до 1,3-1,4 г/г а.с.м. ткани, независимо был ли период осень-зима «холодным» или «тёплым».
Рост содержания воды в тканях меристем почек ели и сосны весной начинался не одновременно. В ели влажность начинала возрастать уже во второй половине марта при отрицательных среднемесячных температурах воздуха, хотя в отдельные дни уже наблюдалась оттепель. У сосны начало роста влажности происходило на две недели позже при наступлении положительных среднемесячных температур воздуха. Весной 2005-2006 гг. (самый холодный год) начало роста влажности отмечено при среднемесячных температурах около минус 2°С (в отдельные дни наблюдались среднесуточные нулевые и положительные температуры воздуха). Темпы роста влажности у сосны были выше и составляли около 0,06 г воды/г а.с.м. ткани в сутки, против 0,04 г воды/г а.с.м. ткани в сутки – роста влажности у ели. В мае перед распусканием хвои влажность в меристемах почек в обеих породах составляла около 3 г воды /г а.с.м. ткани.
Ранее установлено [8], что в почках зимой находятся, по крайней мере, две относительно изолированные фракции воды, одна из которых локализована в клетках меристематических тканей, а другая – в зонах льдовыделения. Место локализации этих зон и соответственно образования льда зависит от строения почек. Различная морфология почек сосны обыкновенной (Pinus silvestris L.) и ели сибирской (Picea obovata L.) обуславливает различные механизмы обезвоживания клеток меристем. Почка ели имеет полость в своем основании ниже зачатков побега и хвои, она непосредственно соединяется с сердцевинной частью побега и является зоной внеорганного льдообразования (рис. 3). Таким образом, в почках ели реализуется механизм внеорганного льдообразования.
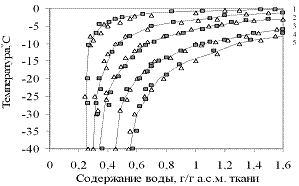
Наглядную характеристику распределения жидкой фазы воды, находящейся при отрицательных температурах в равновесии со льдом, между растворимыми и нерастворимыми компонентами меристематических клеток дают кривые равновесного водосодержания (рис.5).
Анализ кривых показывает, что при снижении температуры до минус 20 °С в меристематических тканях достигается достаточно безопасное содержание воды, равное 0,50 ±0,03 г/г а.с.м. ткани. К этому времени в комплексе клеточных мембран практически не остаётся воды, способной кристаллизоваться и, таким образом, повреждать клеточные мембраны. Тем не менее, в клетках вплоть до температуры около минус 40 °С остаётся жидкая фаза воды, связываемая растворимыми веществами цитоплазмы, причём преимущественно низкомолекулярными веществами. При снижении температуры до минус 40 °С остаточное содержание воды в меристематических тканях снижается до 0,35 ± 0,02 г/г а.с.м. ткани.
Иное строение имеет почка сосны, в которой нет полости, а сердцевинная ткань является продолжением сердцевины побега и направленно связана с меристематическими тканями (рис. 4). Вода, мигрируя в межклеточное пространство, кристаллизуется в нём. Таким образом, здесь реализуется механизм внеклеточного льдообразования. В этом случае значение равновесного водосодержания совпадает с количеством невымораживаемой воды.
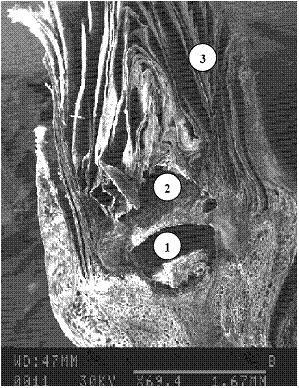
1 – полость в основании почки;
2 – меристематическая ткань;
3 – кроющие чешуи
Рисунок 3 – Микрофотография продольного разреза вегетативной почки ели сибирской
Таким образом, обезвоживание меристематических тканей почек ели и сосны начиналось одновременно в сентябре ещё при положительных температурах воздуха. Следовательно, подготовка меристем почек к состоянию низкотемпературной устойчивости происходит задолго до начала сезонного действия температурного стрессового фактора – снижения среднесуточных температур до минусовых значений. Полученные данные позволяет говорить о том, что меристематические ткани почек ели обыкновенной и сосны обыкновенной обладают наследственно закреплённым механизмом криорезистентности, который реализуется в состоянии глубокого покоя, независимо от того, находится дерево или нет в условиях низкотемпературного стресса.
Высокая скорость снижения водосодержания и набухания меристем у сосны, по-видимому, объясняет достижение минимального водосодержания в меристемах её почек осенью на две недели раньше, чем у ели, несмотря на то, что процесс обезвоживания начинался в обеих породах одновременно. Весной, напротив, начало набухания меристем ели на две недели опережало начало набухания меристем сосны. В то же время следует отметить, что снижение как среднесуточных, так и среднемесячных температур осенью коррелирует со снижением содержания воды в тканях меристем. Весной процесс набухания тканей также коррелирует с изменением температуры воздуха (рис. 6).
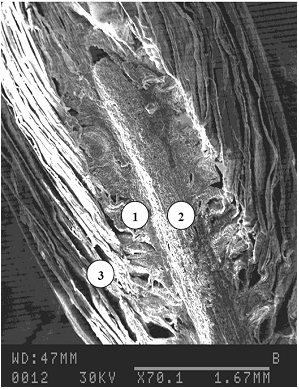
1 - меристематические ткани;
2 - сердцевинная ткань почки;
3 - кроющие чешуи
Рисунок 4 - Микрофотография продольного разреза вегетативной почки сосны обыкновенной
1 - низкомолекулярные соединения; 2 - растворимые вещества цитоплазмы; 3 - меристематические ткани почек ели и сосны; 4 - высокомолекулярные соединения (водорастворимые белки цитоплазмы); 5 - комплекс клеточных мембран; ? - ель; – сосна
Рисунок 5 - Кривые равновесного содержания воды надо льдом в области температур 0 ? минус 40°С
Одинаковое минимальное содержание воды в меристематических тканях почек обеих пород в состоянии низкотемпературной устойчивости, с одной стороны, может свидетельствовать
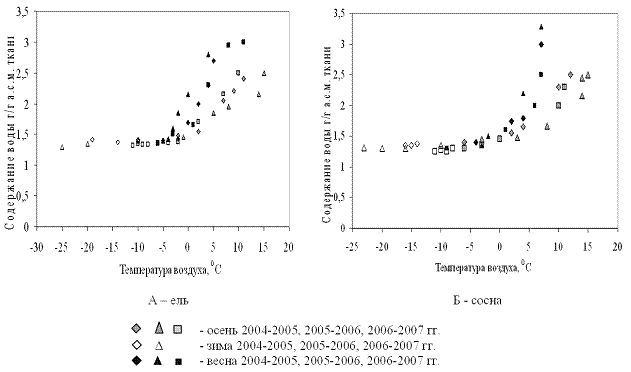
Рисунок 6 - Взаимосвязь влажности меристематических тканей почек ели и сосны и сезонных изменений температуры воздуха
о сходстве механизмов криорезистентности, а с другой, – требует более детального анализа, поскольку отличия в низкотемпературном поведении воды в почках ели и сосны при формировании криозащищённого состояния их меристем, очевидно, обеспечиваются на уровне процессов метаболизма меристематических клеток. Наличие жидкой фазы воды в клетках вплоть до минус 40°С позволяет предположить, что именно поэтому при низких температурах возможно протекание биохимических процессов, проявляющееся в изменении содержания, состава и свойств компонентов клеток.
Библиографический список
- Красавцев, О.А. Отток воды из переохлаждённых зачаточных бутонов / О.А. Красавцев, О.Н. Разнополов, Н.Н. Хвалин // Физиология растений. - 1983. – Т.3. – Вып. 5. – С. 1025.
- Quamme, H.A. Mechanism of supercooling in overvintering peach flover buds / H.A. Quamme // J. Amer. Soc. Horticult. Sci., 1978. - V. 103. – p. 57.
- Sakai, A. Freezing avoidance mechanism of primordial shoots of conifer buds / A. Sakai // Plant and Cell Phisiol. - 1979. – V. 20. – № 7. - p. 1381.
- Агроклиматические ресурсы Красноярского края и Тувинской АССР: справочник. – Л.: Гидрометеоиздат, 1974. – 211 с.
- Алаудинова, Е.В. Сезонные изменения состава и свойств белков и фосфолипидов меристематических тканей почек лиственницы сибирской: дис… на соиск. уч. ст. канд. хим. наук / Е.В. Алаудинова. - Красноярск, 2000. – 145 с.
- Гавриленко, В.Ф. Большой практикум по физиологии растений. Фотосинтез. Дыхание: учеб. пособие / В.Ф. Гавриленко, М.Е. Ладыгина, Л.М. Хандобина. – М.: Высш. школа. – 1975. – 392 с.
- Миронов, П.В.. Переохлаждение и обезвоживание хвойных зачатков в зимующих почках лиственницы сибирской / П.В. Миронов, Э.Д. Левин // Физиология растений. - 1985. – Т. 32. – Вып. 4. – С. 695-700.
- Миронов, П.В. Низкотемпературная устойчивость живых тканей хвойных: монография [Текст] / П.В. Миронов, Е.В. Алаудинова, С.М. Репях. – Красноярск: СибГТУ, 2001. – 221 с
|


