|
"Хвойные бореальной зоны" 2007г.,№2-3, с.
ПОПУЛЯЦИОННО-ГЕОГРАФИЧЕСКАЯ ДИФФЕРЕНЦИАЦИЯ ЕЛИВ ЗАПАДНОЙ СИБИРИ
Попов П.П.
Институт проблем освоения Севера СО РАН
625003, Россия. Тюмень-3, а/я 2774,
ipospopov@mail.ru
Изучение популяций проведено с использованием кластерного и дискриминантного анализов по двум показателям, характеризующим форму семенных чешуй: коэффициентам сужения (Cn) и вытянутости (Cp) их верхней части [Попов, 1999]. Материалом для анализа являются популяционные выборки (27 шт., 5000 деревьев) шишек ели на территории от западных районов Тюменской и восточных (районов) Свердловской областей до Енисея (Красноярск – Игарка). Крайними вариантами популяций для дискриминантного анализа взяли выборку "типичной" ели европейской с Карпат и выборку шишек «типичной» ели сибирской из района Олекминская (Саха (Якутия) и Иркутской области (Витимский заповедник). Ель в Западной Сибири в феногенетическом отношении близка к "типичной" ели сибирской из Восточной Сибири. Она характеризуется сравнительно небольшим популяционным разнообразием, изменяющимся с юго-запада на северо-восток; и представлена в основном двумя группами: западной и юго-западной (группой) до Оби–Иртыша и северо-восточной и восточной (группой). Такая дифференциация популяций обусловлена, вероятно, генетическим влиянием ели с Урала и Предуралья.
Population study ful fil to Klaster and Discriminant analyses an two index forms of scale: coefficient of narrowing (Cn) and coefficient of projection (Cp) [Popov, 1999]. Material of analyses to be populations sample (27s., 5000 trees) cones fir Tyumen and Sverdlovsk area to river Enisey (Krasnoyarsk – Igarka). Extreme variants of population to discriminant analyses use sample «typical» Picea abies (L.) Karst. in Karpatyca and sample cones «typical» Picea obovata Ledeb. in Olekminsk (Saha)Yakutya) and Irkutsk area (Vitimsky reservation). Picea (fir) in West Sibirian to phene near to «typical» Picea sibirian (P. obovata) in Ost Sibirian. She to have small of population variation, changeable with west to ost: two group – westen group (to river Ob – Irtysh) and osten group. To such differentiation of populations to be conditioned, in all probability, genetic influence fir the Ural and West Ural.
Введение
Еловые леса в Западной Сибири занимают площадь около 15 млн. га. Они располагаются на обширных пространствах от подзоны южной тайги до почти северной границы леса. В пределах южной и отчасти средней тайги древостои обычно относятся к IV-III классам бонитета, а на остальной территории, севернее, примерно, 61-62? ельники характеризуются V классом бонитета, т. е. в целом они обладают сравнительно невысокой производительностью [Цепляев, 1961; Крылов, Крылов, 1969; Атлас, 1973]. Однако еловые леса здесь играют важную социально-экологическую роль. Предшествующие исследования показали, что ель в Западной Сибири в географическом и эволюционно-генетическом отношениях «связана» с елью, произрастающей на Урале и в Предуралье [Кеппен, 1885; Правдин, 1975; Морозов, 1976 и др.]. Научный и лесоводственный интерес представляет изучение популяционно-географической дифференциации ели на территории Западной Сибири относительно "типичных" популяций ели европейской (Piceaabies (L.) Karst.) и ели сибирской (Piceaobovata Ledeb.). Это и составляет цель работы.
Экспериментальная часть
Изучение популяций проведено с использованием кластерного и дискриминантного анализов [Боровиков, 1998; Гашев, 1998] по двум признакам (пока
зателям), характеризующим форму семенных чешуй: коэффициентам сужения (Cn) и вытянутости (Cp) их верхней части [Попов, 1999]. Изменчивость формы семенных чешуй елей серии Obovatae Bobr. [Бобров, 1971] в большой степени определяется генотипом особей. Поэтому, изучая изменчивость популяций по форме семенных чешуй, мы, по-существу, выявляем их наследственную дифференциацию, в этом и состоит главная особенность подобных исследований.
Исходными материалами для анализа послужили популяционные выборки шишек ели на территории от западных районов Тюменской и восточных (районов) Свердловской областей до Енисея (Красноярск – Игарка). Крайними ("эталонными") вариантами популяций для дискриминантного анализа взяли выборки из Закарпатской области Украины (г. Рахов) с территории Карпатского биосферного заповедника (370 особей) и Ивано-Франковской обл. (100
особей), представляющие "типичную" ель европейскую. В качестве "типичных" представителей ели сибирской использовали выборки шишек из окрестностей г. Олекминска (Саха (Якутия) и с территории Витимского биосферного заповедника в северо-восточной части Иркутской области. В последнем случае объединенный образец включает 275 (125+150) особей. Главными критериями при выборе "эталонных" (крайних по видам ели) популяций было предположение их "видовой чистоты", т.е. отсутствие генетического влияния друг на друга и большая величина выборок.
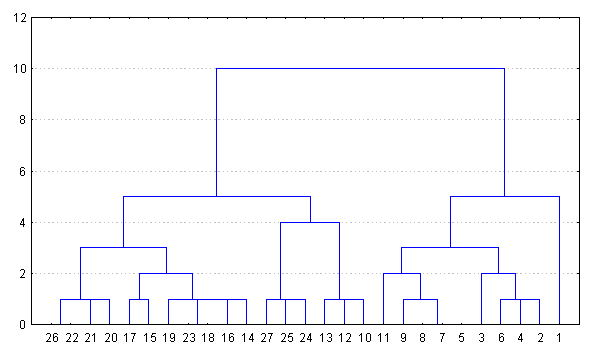
Пункты
Рисунок 1 - Дендрограмма (Chebychev distance metric), составленная по средним значениям коэффициентов вытянутости (Cp) и сужения (Cn) формы семенной чешуи ели в Западной Сибири. Пункты: 1 - Тюмень, 2 - Вагай, 3 - Междуреченский, 4 - Зеленоборск, 5 - Уват, 6 - Чембакчино, 7 - Саранпауль, 8 - Кыштовка, 9 - Аламбай, 10 - Томск, 11 - Красноярск, 12 - Енисейск, 13 - Подкаменная Тунгуска, 14 - Угут, 15 - Нижневартовск, 16 - Ханты-Мансийск, 17 - Когалым, 18 - Октябрьский, 19 - Полноват, 20 - Овгорт, 21 - Салехард, 22 - Надым, 23 - Новый Уренгой, 24 - Толька, 25 - Красноселькуп, 26 - Сидоровск, 27 - Игарка
Категории или формы особей ели в популяциях принимаются следующие: из района Карпат – P.e. (как Picea europaea), Витима и Олекмы – P.s. (как Picea sibirica), для "промежуточной ели" – P.m. (как Picea medioxima). Все полученные результаты высокодостоверны, поскольку число особей в выборках достаточно велико. Квадрат расстояния Махаланобиса (Squared Mahalanobis Distances – SMD) между выборками из района Карпат и Витима и Олекмы составляет 104.23. В качестве "промежуточной ели" взяли выборки из районов Никольска (Вологодская обл.) и Коноши (Архангельская обл.). В этих выборках, включающих по 130 особей, количество форм P.e. и P.s. оказалось практически одинаковым. И с этой точки зрения данные выборки представляют собой действительно промежуточную популяцию. SMD между P.e. и P.m. (объединенный образец, включающий 260 особей) равен – 20,5, между P.s. и P.m. – 16,6, а между P.e. и P.s. в этом случае – 66,3.
Если брать выборки только из Закарпатья и района Олекмы, то SMD равен 117.44; из Закарпатья, Витима и Олекмы – 118.55, т. е. квадрат расстояния Махаланобиса оказывается даже несколько большим, чем при сравнении ели сибирской (из района Витима и Олекмы) с двумя выборками с Карпат (Рахов и Ивано-Франковск). Но во всех случаях сравниваемые выборки "типичных" елей европейской и сибирской весьма существенно различаются и полигоны распределения особей (на Scatterplot of canonical scores) по показателям формы семенных чешуй не перекрываются (трансгрессии нет).
Результаты и их обсуждение
Исследования ели на территории Тюменской области [Попов, 1987; Муканова, Попов, 2001] позволили установить определенные отличия популяций в западных районах от популяций, располагающихся далее к востоку. Западные популяции отличаются, прежде всего, тем, что в них встречаются особи, имеющие заметно выраженную угловатость (заостренность) семенных чешуй и, в связи с этим, средний показатель коэффициента сужения (Cn) оказывается несколько меньше, а коэффициент вытянутости (Cp) верхней части чешуй, напротив, больше, чем в северных и восточных районах.
Для всей территории Западной Сибири по средним показателям (Cn, Cp) формы семенных чешуй составлена дендрограмма (рисунок 1). Общий кластер четко разделяется на два (кластера) с величиной дистанции Чебышева (Chebychev distsnces metric – CDM), равной 5. Каждый из них в свою очередь, разделяется еще на два с величиной CDM – 3–4.
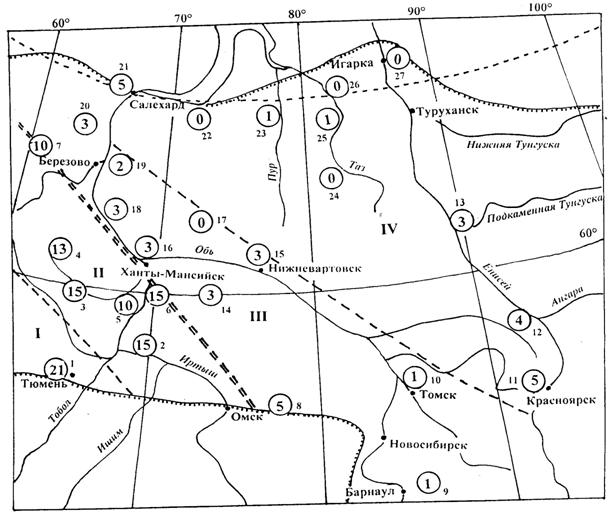
Рисунок 2 - Частота особей фенотипа P.m. (цифры в кружках) в популяциях ели сибирской на территории Западной Сибири по результатам дискриминантного анализа. Цифры рядом с кружками – № пунктов сбора материалов
Условно первый кластер составляют выборки из района Тюмени (Велижаны, Янтык, Леваши, Успенка, Тугулым, Юшала). Ель здесь ближе и к ели, произрастающей к северо-востоку от Тюмени (район II на рисунке 2). Последняя также составляет отдельный кластер, который «объединяется» с тюменской группой популяций. Восточная граница этой группы популяций проходит примерно от Саранпауля, западнее Оби, к верхнему течению р. Демьянки и далее до границы ареала. В целом она имеет общее направление северо-запад – юго-восток, и удивительно точно совпадает с западной границей ареала Larixsibirica L. и восточной (границей) L. SukaczewiiDyl. [Коропачинской, Милютин, 2006]. Еще далее к северо-востоку выделяются еще две группы популяций, занимающие районы III и IV.
При разделении особей на две категории (P.e. и P.s.) с помощью дискриминантного анализа оказывается (таблица), что в популяциях западной и юго-западной части региона единично (до 5 %) встречаются особи, морфотип которых ближе к ели европейской (P.e.), чем к сибирской (P.s.). Сюда относятся: район Тюмени (особей формы P.e. 1– 4,7 %), Вагая (до 2 %), Верхней Конды, Чембакчиной (Ханты-Мансийское прииртышье) (по 1 %). Вся остальная часть особей в популяциях (т. е. 95 % и более) соответствует морфотипу P.s.
При разделении особей на три категории (P.e., P.m., P.s.) морфотипа P.e. не оказывается ни в одной выборке, но во всех районах имеется разное количество (1-30 %) особей морфотипа P.m. (таблица). Географическое расположение популяций по количеству особей типа P.m. позволяет разделить их, по крайней мере, на 4 района (рисунок 2). В районе I количество их составляет в среднем около 20(15-30) % (особей морфотипа P.s., соответственно, 80 %). В районе II особей морфотипа P.m. 10-15 %, в районе III их всего 2-5 %, в районе IV их практически нет (0-1 %). Довольно неожиданным является повышенная встречаемость (3–5 %) таких особей в приенисейских популяциях ели, что можно объяснить отепляющим действием большой реки, при котором их больше выживает. Как при кластерном, так при дискриминантном анализах получается довольно четкое разделение всех популяций на две группы. В первую входят районы I и II, во вторую – III и IV.
Таблица 1 - Фенотипическая структура популяций ели в разных районах Западной Сибири
№ на рис. 2 |
Группы и районы популяций.
Пункты |
Число особей, шт. |
Частота фенотипов, % |
при 2-классной градации |
при 3-классной градации |
P.e. |
P.s. |
P.e. |
P.m. |
P.s. |
1
1а
1б
1в
1г
1д
2
3
4
5
6
7
8
21
20
19
18
16
14
15
11
12
13
10
9
17
22
23
24
25
26
27 |
Группа I. Район I
Тюмень (итог):
Леваши
Успенка
Велижаны
Янтык
Юшала и Тугулым
Группа I. Район II
Вагай
Междуреченский
Зеленоборск
Уват
Чембакчина
Саранпауль
Группа II. Район III
Кыштовка
Салехард
Овгорт
Полноват
Октябрьский
Ханты-Мансийск
Угут
Нижневартовск
Красноярск
Енисейск
Подкамен. Тунгуска
Группа II. Район IV
Томск
Аламбай
Когалым
Надым
Новый Уренгой
Толька
Красноселькуп
Сидоровск
Игарка |
1042
235
165
175
167
300
400
250
212
100
150
90
110
195
285
100
100
200
110
200
140
130
170
150
100
110
117
100
220
100
40
210 |
2
5
3
1
2
1
2
-
1
1
1
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
- |
98
95
97
99
98
99
98
100
99
99
99
100
100
100
100
100
100
100
100
100
100
100
100
100
100
100
100
100
100
100
100
100 |
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
- |
21
24
30
17
23
15
15
15
13
10
15
10
5
5
3
2
3
3
3
3
5
4
3
1
1
-
-
1
-
1
-
- |
79
76
70
83
77
85
85
85
87
90
85
90
95
95
97
98
97
97
97
97
95
96
97
99
99
100
100
99
100
99
100
100 |
Примечание. Фенотипы: P.e. - ели европейской, P.s. - ели сибирской, P.m. - промежуточной ели
Наличие в популяциях особей типа P.m. можно объяснить влиянием генома ели европейской. В данном случае особи формы P.m., скорее всего, – результат гибридизации "чистой" ели сибирской и близкой к "типичной гибридной" (ели), произрастающей в Предуралье и на Урале [Попов, 1996]. Относительная численность таких типов (P.m.) в популяциях может отражать уровень или степень влияния гибридизации на структуру популяций. Преобладающая часть особей относится к морфотипу P.s. К этому же типу относятся и все популяционные выборки по средним показателям Cn и Cp. Следует отметить и то, что особи, относящиеся к морфотипу P.e. (при 2-классной градации) или – P.m. (при 3-классной градации), занимают пограничное положение, приближаясь к морфотипу P.s.
В соответствии с принятым разделением популяций имеется четкая их дифференциация по SMD, особенно от «типичной» ели европейской. Величина квадрата дистанции Махаланобиса (SMD) от ели европейской составляет от 81 до 118. В среднем по районам она равна 81, 90,5, 104, 113. Дистанция от «типичной» ели сибирской для популяций в районах II, III и IV составляет менее 1 (0,09-0,72), и только для района I она равна 1,66. Отсюда видно, как далеки западносибирские популяции ели от «типичной» ели европейской и как они близки к «типичной» ели сибирской.
Следующим важным показателем дифференциации западносибирских популяций и принадлежности их к ели сибирской является критерий (коэффициент) вероятности (Posterior Probabilities – P.P.). По районам популяций он в среднем составляет: 0,68, 0,82, 0,94, 0,97 (или, соответственно, 68, 82, 94, 97 %). Таким образом, несмотря на значительную вариацию этого показателя, западносибирские популяции относятся к ели сибирской. Причем, в районах III и IV они особенно близки к ели из Восточной Сибири; по уровню вероятности их отличие составляет только 3-6 %. В районах I и II отличие значительно больше (32 и 18 % соответственно).
Таким образом, ель, произрастающая в Западной Сибири, заметно разделяется, по крайней мере, на 4 района, объединяемых в 2 группы популяций. В феногенетическом отношении она близка к "типичной" ели сибирской из Восточной Сибири. Этот вывод не противоречит сложившимся представлениям [Правдин, 1975; Соколов и др., 1977; Бобров, 1978 и др.], но в данном исследовании он получает вполне объективное подтверждение.
Библиографический список
- Атлас лесов СССР. - М.: ГУГК при СМ СССР, 1973. - 222 с.
- Бобров, Е.Г. История и систематика рода Picea A. Dietr. / Е.Г.Бобров // Новости систематики высших растений. 1971. - Вып. 7. - С. 5-40.
- Бобров, Е.Г. Лесообразующие хвойные СССР [текст] / Е.Г. Бобров. - Л.: Наука. Ленингр. отд-е, 1978. -188 с.
- Боровиков, В.П. Популярное введение в программу STATISTICA [текст] / В.П.Боровиков .- М.: КомпьютерПресс, 1998.- 267 с.
- Гашев, С.Н. Статистический анализ для биологов (Пакет программ "STATAN") [текст] / С.Н. Гашев.-Тюмень: Изд-во ТюмГУ, 1998. - 51 с.
- Кеппен, Ф. Географическое распространение хвойных деревьев в Европейской России и на Кавказе [текст] / Ф.Кеппен.- СПб, 1885. - 634 с.
- Коропачинский, И.Ю. Естественная гибридизация древесных растений [текст] / И.Ю. Коропачинский, Л.И. Милютин - Новосибирск: Академиздательство «Гео», 2006. - С. 223.
- Крылов, Г.В. Леса Западной Сибири. / Г.В. Крылов, А.Г. Крылов // Леса СССР: Леса Урала, Сибири и Дальнего Востока. -М.: Наука, 1969. - Т.4. - С.157-247.
- Морозов, Г.П. Фенотипическая структура популяций ели обыкновенной и сибирской / Г.П. Морозов // Лесоведение. 1976. - № 5. - С. 22-29.
- Муканова, А.А. Фенотипическая структура популяций ели сибирской на территории Тюменской области / А.А.Муканова, П.П. Попов // Вестник экологии, лесоведения и ландшафтоведения. Тюмень: ИПОС СО РАН, 2001. - Вып. 2. - С. 97-104
- Попов, П.П. Изменчивость генеративных органов ели сибирской в Тюменской области / П.П. Попов // Лесоведение. - 1987. - № 3. - С. 27-32.
- Попов, П.П. Гибридная ель на северо-востоке Европы / П.П. Попов // Лесоведение. - 1996. - № 2. - С. 62-72.
- Попов, П.П. Ель на востоке Европы и в Западной Сибири [текст] / П.П. Попов.- Новосибирск: Наука, 1999.- 169 с.
- Правдин, Л.Ф. Ель европейская и ель сибирская в СССР [текст] / Л.Ф.Правдин. - М.: Наука, 1975.- 176 с.
- Соколов, С.Я. Ареалы деревьев и кустарников СССР [текст] / С.Я. Соколов, О.С. Связева, В.А. Кубли.- Л.: Наука, 1977.- 163 с.
- Цепляев, В.П. Леса СССР [текст] / В.П. Цепляев.- М.: Сельхозиздат, 1961.- 216 с.
|


