|
Влияние пожаров на интенсивность дыхания ствола сосны обыкновенной (Pinus silvestrisL.)
Масягина О.В., Прокушкин С.Г., Иванова Г.А.
Институт леса им. В.Н. Сукачева СО РАН, Красноярск
Работа выполнена при финансовой поддержке российско-американского проекта (№ 076) и гранта РФФИ № 03-04-48037.
Определена ответная реакция ствола сосны обыкновенной (PinussilvestrisL.) на термическое воздействие при низовом пожаре. Дыхание ствола в зависимости от жизненного состояния деревьев сосны определяли insitu в сосняках Нижнего Приангарья в июле-августе. Выявлена интенсивность дыхания ствола до начала выжигания и через сутки после пожара. В большинстве случаев интенсивность дыхания ствола у деревьев различного жизненного состояния достоверно отличалась до и после выжигания. Анализ показал, что до проведения выжигания среднее дыхание ствола в группе здоровых деревьев в 1.5-1.6 раза выше, чем у ослабленных и сильно ослабленных деревьев. Различий в дыхании ствола между ослабленными и сильно ослабленными деревьями не обнаружено. На следующий день после пожара отмечено снижение интенсивности дыхания ствола в группах здоровых и сильно ослабленных деревьев, в то время как в группе ослабленных деревьев дыхание возрастало.
Response of Pinus silvestris L. on thermal impact at ground fire has been studied. Stem respiration subject to vital status of pine trees was determined in situ in pine stands of Nijnee Priangarie in July-August. Stem respiration rate before and after ignition have been revealed. In most cases stem respiration rate of different vital tree status reliably differed before and after fire. Statistical analysis showed that before fire, average stem respiration rate of healthy trees was 1.5-1.6 times higher than that of weakened or highly weakened trees. In addition, there were no differences in stem respiration found between weakened or highly weakened trees. On the next day after ground fire, decreasing of stem respiration rate of healthy and highly weakened trees was observed whereas stem respiration rate of weakened trees was increasing.
Введение
В лесных экосистемах значительная часть всего углерода находится в древесине, и по оценкам в фитомассе лесов России ежегодно депонируется 16% от общего количества накопившегося углерода лесов, что составляет около 207*109 т С [1]. В связи с тем, что лесные экосистемы балансируют на грани между состоянием «источника СО2 в атмосферу» и «стока углерода» [2] назрела необходимость проверки точности моделей, оценивающих бюджет углерода лесов и таким образом расширить наши знания о процессах, лежащих в основе эмиссии СО2 древостоем.
В расходной части углекислотного газообмена насаждения выделение углекислоты стволами древесных растений составляет значительную величину, так как биомасса стволов занимает существенную часть древостоя. Интенсивность дыхания стволов варьирует в зависимости от породы и возраста дерева, сезона и других биогенных и абиогенных факторов [3, 4]. Дыхание может потреблять от 11% до70% углерода, фиксированного растениями при фотосинтезе [5, 6], и по некоторым оценкам вклад дыхания ствола в общий бюджет углерода надземной части спелого древостоя составляет от 13% [7] до 42% [3, 5]. Следовательно, вклад древесной ткани в расходную статью бюджета углерода надземной части древостоя составляет по крайней мере 25%, и изменения в этом компоненте в связи со стрессовыми воздействиями (пожары, вырубки, техногенные нагрузки и т.д.) могут привести к значительным изменениям в бюджете углерода лесных экосистем. Пожары оказывают значительное влияние на бюджет углерода (на его пулы и потоки), так как каждый год повреждается 2-3 млн. га лесов [8, 9]. Наибольший эффект наблюдается от верховых пожаров, но низовые пожары, даже при сохранившемся древостое, способны влиять на СО2-газообмен лесной экосистемы, в том числе и на дыхание самого древостоя. Несмотря на всю значимость дыхания древесной ткани для бюджета углерода экосистемы, данных по влиянию пожаров на дыхание ствола древесных растений недостаточно.
Кроме того, в одновозрастном насаждении может наблюдаться большое разнообразие роста и развития между отдельными древесными растениями, которые с помощью ряда морфометрических показателей объединяют в соответствующие группы или классы роста. Древесные растения каждого класса роста характеризуются различной скоростью физиологических процессов, в том числе и дыхания ствола. Однако насколько значительны эти различия до сих пор остается слабо изученными. Вместе с тем исследование особенностей дыхания ствола деревьев различного роста и развития не только дополняет морфофизиологическую характеристику насаждения в целом, но и может быть критерием их физиологического состояния в каждой из выделенных групп. Кроме того, подобные сведения важны при расчете общего выделения углекислого газа поверхностью стволов, так как это позволит более точно оценивать расходную статью баланса СО2-газообмена в рамках всего насаждения [3].
В связи с этим, целью данной работы было определение кратковременной реакции дыхания ствола на термическое воздействие, возникающее при пожаре, так как это позволит понять ответный отклик всего древостоя на изменения окружающей среды, связанные с изменением экологических условий.
Экспериментальная часть
Исследования, являющиеся частью совместного российско-американского проекта «The Russian FIRE BEAR» (Fire Effects in the Boreal Eurasia Region), проводили в сосняках Нижнего Приангарья Красноярского края (58о42? с.ш., 98о25? в.д., абс. высота н.у.м. 200-300 м, Хребтовский лесхоз, 55 км от Богучанской ГЭС вниз по течению р. Ангара) в июле-августе 2003 г. Цель данного проекта – моделирование поведения лесных пожаров разной интенсивности и их влияние на экосистему. Территория относится к подзоне южной тайги. Детальное описание экспериментального дизайна дано у Ивановой [9]. Были подобраны два участка (3 и 5) площадью 200 х 200 м для проведения последующего выжигания, где горение распространялось фронтальной кромкой по ветру. Пожары были квалифицированы как низовые низкой интенсивности, но при этом скорость распространения кромки огня была выше на 5-м участке (таблица 1). Характеристика сосняков приспевающих разновозрастных, где проводились исследования, приведена в таблица 2.
Почва 3-го участка классифицируется как неглубокоподзолистая типичная песчаная, а почва 5-го участка – иллювиально-железистый песчаный подзол [10] характеризуется высокой кислотностью, низким содержанием гумуса и низкой трофностью. Лесная подстилка бурая, средней степени разложения. Климат района резко континентальный, среднемесячная температура января от -22.0оС до -33.1оС, а июля от +17.8оС до +18.4оС, количество осадков 320-380 мм в год [9].
Дыхание ствола деревьев сосны (PinussilvestrisL.) определяли insitu в сосняках на участках 3 и 5 (таблица 2 и 3) в конце июля-начале августа 2003 г. при помощи ИК-газоанализатора Li-Cor 6200 (LI-COR, США) в дневное время суток (с 12:30 до 17:30). Дыхание ствола определялось в зависимости от жизненного состояния деревьев, которое определяли по методике Алексеева [11].
Таблица 1 - Количественные характеристики поведения огня на экспериментальных участках при низовом пожаре*
№ участка |
Дата эксперимента |
Количество сгорающей биомассы, кг/м2 |
Глубина прогорания, см |
Скорость распространения кромки огня, м/мин |
Температура на поверхности мохово-лишайникового покрова, оС |
Интенсивность огня, кВт/м |
3 |
01.08.2003 |
1.6 |
3.5 |
1.0 |
570 |
<100 |
5 |
31.07.2003 |
1.3 |
3.1 |
1.2 |
600 |
<200 |
* - по данным MacRae [29].
Таблица 2 - Характеристика древостоев на пробных площадях
№ п.п. |
Тип леса и местоположение |
Состав |
Полнота |
Нср, м |
Дср, см |
Подрост, 103 экз./га |
Число стволов, шт/га |
3 |
Сосняк ольховниково-бруснично-зеленомошный |
10С+Л |
0.6 |
18-22 |
18-32 |
20 |
250 |
5 |
Сосняк бруснично-зеленомошный |
10С+Л,ед.Ос |
0.8 |
18-22 |
18-20 |
27 |
270 |
Выделено три группы деревьев: здоровые, ослабленные и сильно ослабленные. Каждая группа была представлена тремя биологическими повторностями. Измерение интенсивности дыхания ствола проводили до выжигания и на следующий день после выжигания.
Интенсивность дыхания определяли на интактных стволах во избежание возникновения индукции дыхания или травматического дыхания [12], так как это явление может привести к завышенным результатам и сложности сопоставления данных с результатами других исследователей. Перед измерением дыхания кору дерева зачищали на высоте 1.3 м до образования ровной поверхности. Затем устанавливали темновую камеру (модель 6000-09), которая исключает возможность ассимиляции углекислоты корой внутри камеры. Объем камеры составляет 962 см3, а площадь сечения 78.5 см2. Стык камеры с корой тщательно изолировался герметиком. Количество повторностей измерения дыхания ствола варьировало от 3 до 6. Температура воздуха, замеряемая во время замера дыхания, варьировала в пределах от 20оС до 30оС.
Таблица 3 - Характеристика опытных деревьев сосны обыкновенной до выжигания
№ дерева |
Жизненное состояние |
Д,
см |
Высота,
м |
Возраст на 2003 год, лет |
Охвоенность кроны |
Протяженность кроны, м |
3-й участок |
336 |
I |
16.0 |
20.0 |
55 |
средняя |
6.0 |
165 |
22.0 |
23.0 |
55 |
средняя |
4.0 |
335 |
24.0 |
23.0 |
55 |
сильная |
4.0 |
320 |
II |
18.0 |
20.0 |
55 |
слабая |
3.0 |
191 |
14.0 |
17.0 |
55 |
слабая |
5.0 |
347 |
19.0 |
22.0 |
55 |
слабая |
5.0 |
326 |
III |
13.0 |
17.0 |
55 |
очень слабая |
4.0 |
328 |
18.0 |
23.0 |
55 |
слабая |
4.0 |
108 |
15.0 |
18.0 |
55 |
очень слабая |
2.0 |
5-й участок |
395 |
I |
24.0 |
22.0 |
55 |
сильная |
8.0 |
161 |
17.0 |
21.0 |
55 |
сильная |
7.5 |
340 |
17.0 |
20.0 |
55 |
сильная |
5.0 |
106 |
23.0 |
25.0 |
55 |
средняя |
10.0 |
187 |
II |
12.5 |
18.0 |
55 |
средняя |
6.0 |
312 |
15.0 |
18.0 |
55 |
средняя |
5.0 |
150 |
15.0 |
18.5 |
55 |
средняя |
6.0 |
327* |
III |
14.0 |
20.0 |
55 |
слабая |
2.5 |
997 |
22.0 |
23.0 |
55 |
слабая |
7.0 |
302 |
14.0 |
18.0 |
55 |
слабая |
2.5 |
Примечание: I - здоровое, II - ослабленное, III - сильно ослабленное, *во время пожара луб у корневой шейки дерева поврежден на 50-70%.
Расчет дыхания производился на поверхность ствола и выражался в mмолях СО2/(м2 с). Перед проведением выжигания зачищенные участки коры защищали от воздействия огня стекловолокном и фольгой. Через сутки после выжигания дыхание замеряли на тех же участках ствола, что и до выжигания.
Статистический анализ полученных результатов проводили с помощью процедуры GLM пакета Statistica 7.0 (StatSoft, США).
Результаты и обсуждение
До начала выжигания интенсивность дыхания ствола сосны обыкновенной составила от 1.0 до 11.3 mмоль СО2/(м2 с) (таблица 4), что сопоставимо с величинами дыхания, полученными различными авторами. Так, интенсивности дыхания ствола приведенные в книге «Рост и газообмен СО2 у лесных растений» [13] для PinussilvestrisL. (100-летний древостой) за июнь-август имели сходные результаты с полученными нами пределами варьирования - от 0.08 до 0.5 мг СО2/(м2 с) (или от 1.8 до 10.5 mмоль СО2/(м2с)). При этом возраст древостоя оказался близким с нашим. Однако, полученные значения интенсивности дыхания ствола оказались несколько выше значений, полученных различными авторами для различных видов сосны, где дыхание ствола варьировало от 0.1 до 5.1 mмоль СО2/(м2 с) [3, 14, 15].
Однако в некоторых случаях проводить сравнение с данными по стволовому дыханию сосны сложно, так как в них расчет дыхания ведется на объем заболони, что связано с положительной корреляцией между выделением углекислоты с поверхности ствола и объемом заболони [14, 16].
Исследования по определению объема заболони нами не проводилась, но в некоторых работах отмечается, что в среднем мощность заболони не превышает 5 мм. В частности, по данным Han и Suzaki [17] слой интенсивно дышащих тканей на боковой поверхности ствола составлял примерно всего 5 мм, что является незначительной частью объема ствола взрослого дерева (до 5% на высоте ствола 1.3 м; [14]), клетки этого слоя выделяют до 95% всей углекислоты, образующейся при дыхании этой части дерева.
Таблица 4 - Интенсивность дыхания стволов сосны до и после проведения выжигания, mмоль СО2/(м2 с)
№ дерева |
Жизненное состояние |
До выжигания |
После выжигания |
Средняя интенсивность дыхания |
Среднее дыхание ствола внутри группы |
Средняя интенсивность дыхания |
Среднее дыхание ствола внутри группы |
3-й участок |
336 |
I |
6.3±0.4 |
8.4±0.8 |
5.0±0.4 |
4.2±0.6 |
165 |
11.3±0.6 |
1.7±0.2 |
335 |
7.6±0.5 |
5.6±0.3 |
320 |
II |
4.3±0.6 |
5.7±0.7 |
11.1±0.7 |
7.9±1.2 |
191 |
4.6±0.1 |
9.0±0.8 |
347 |
8.0±0.5 |
3.2±0.3 |
326 |
III |
4.4±0.4 |
6.1±1.0 |
2.3±0.1 |
7.1±2.0 |
328 |
10.3±0.8 |
14.4±0.6 |
108 |
4.3±0.4 |
4.47±0.18 |
5-й участок |
106 |
I |
3.6±0.4 |
5.6±2.0 |
8.0±0.7 |
6.5±0.6 |
161 |
10.1±0.5 |
5.6±0.5 |
340 |
6.4±0.8 |
5.8±0.4 |
395 |
2.2±0.2 |
6.4±0.3 |
150 |
II |
5.0±0.4 |
5.0±0.9 |
5.3±0.3 |
6.2±0.6 |
187 |
3.7±0.7 |
6.2±0.2 |
312 |
6.3±0.5 |
7.0±0.6 |
302 |
III |
6.8±0.8 |
4.4±2.2 |
4.6±0.5 |
3.5±0.7 |
327 |
1.0±0.1 |
2.8±0.1 |
997 |
5.4±0.5 |
3.2±0.3 |
Примечание: I - здоровое, II - ослабленное, III - сильно ослабленное. Приведены средние по интенсивности дыхания ствола для отдельных деревьев и внутри группы по жизненному состоянию и стандартная ошибка среднего.
Исходя из этого, мы рассчитали примерные величины дыхания в расчете на объем заболони. Как следует из таблицы 4, дыхание ствола до пожара варьирует от 1.0 mмоль СО2/(м2 с) (дерево № 327, участок 5) до 11.3 mмоль СО2/(м2 с) (дерево № 165, участок 3). Согласно данным Ryan [14], радиус заболони для этих деревьев приблизительно составляет 35 мм = 0.0035 м (для дерева № 327) и 55 мм = 0.0055 м (для дерева № 165). Следовательно интенсивность дыхания в пересчете на объем заболони под камерой будет варьировать от 0.0035 (дерево № 327) до 0.062 (дерево № 165) mмоль СО2/(м3 с). Полученный интервал дыхания сосны оказался гораздо ниже по сравнению с данными Lavigne [4], где дыхание сосны составило в среднем от 20 до 100 mмоль СО2/(м3 с). Дыхание ствола сосны (Pinusponderosa) по данным Law [18] также выше и составляет от 2.5 до 19.5 mмоль СО2/(м3 с) при радиусе заболони от 70 до 100 мм.
Интенсивность дыхания ствола у отдельных деревьев сосны достоверно отличалась до и после выжигания на 3-м участке (за исключением дерева № 108) и на 5-м участке (за исключением деревьев № 150, 312 и 340). При этом анализ различий у групп деревьев различного жизненного состояния до и после выжигания показал достоверные различия на 3-м участке только у группы здоровых деревьев, где интенсивность дыхания ствола до выжигания была 8.4 µмоль СО2/(м2 с), а после выжигания снизилась и составила 4.0 µмоль СО2/(м2 с), (t=4.4, p=0.000, табл. 4). На участке 5 различия в дыхании отмечены только у группы ослабленных деревьев, где до выжигания оно составляло 5.0 µмоль СО2/(м2 с), а после - 6.2 µмоль СО2/(м2 с) (t=-2.2, p=0.04, таблица 4).
Анализ, проведенный внутри каждой группы деревьев показал, что дыхание ствола внутри группы также сильно и статистически достоверно отличается друг от друга. Причины такого варьирования могут быть разными. Основными из них являются различия в количестве живых клеток в годичном кольце, продуцируемых на протяженности всего ствола и в неоднородности их распределения в стволе [19], а также транспорте и запасах углеводов в стволе [20, 21]. Кроме того, вместе с током веществ в направлении кроны может переноситься углекислота, выделенная стволом [22, 23] и т.д. В связи с вышеизложенным, те деревья, которые достоверно отличались по интенсивности дыхания внутри группы до начала выжигания, были исключены из дальнейших расчетов по выявлению влияния пожаров на дыхание ствола. Деревья, которые остались после исключения отличающихся по дыханию стволов по методу исключения сомнительных вариант, описанного Лакиным [24], были сведены в таблица 5 и дальнейший анализ проводился уже на основе этих данных. Многие авторы обнаруживают тесную связь дыхания ствола с микроклиматическими факторами (температурой воздуха, температурой ствола, влажностью воздуха и почвы и т.д.) [3, 25, 26, 27, 28]. Очевидно, ведущим фактором в суточной динамике дыхания ствола являются изменения температуры ствола, которые сказываются на скорости протекания процессов метаболизма [13], а также влияют на физический процесс диффузии СО2 из ствола. К сожалению, в данной работе было невозможно оценить температурную реакцию дыхания ствола, так как диапазон изученных температур воздуха не превышал 2-3оС (таблица 5). При пожаре же действие высоких температур до 600оС было кратковременным.
На основании этих данных отмечены различия в дыхании ствола сосны до и после выжигания во всех взятых нами группах на обоих участках, за исключением сильноослабленных деревьев на 3-м участке (рисунок 1).
У здоровых деревьев на обоих участках отмечается достоверное снижение дыхания ствола в 1.3 раза – на 3-м и в 1.4 - на 5-м участке. У ослабленных деревьев на обоих участках наблюдается сходная реакция – достоверное усиление интенсивности дыхания на следующий день после пожара в 2 раза на 3-м участке и в 1.2 раза на 5-м участке. В то время как, у сильно ослабленных деревьев наблюдается тенденция к снижению дыхания ствола, которое на 3-м участке – недостоверно, а на 5-м участке дыхание ствола после выжигания достоверно ниже в 1.7 раза (рис. 1).
Колебания температуры в стволе связаны также с его объемом, объемом и составом газовой фазы в древесине, что может вызвать либо избыточное выделение, либо, наоборот, поглощение углекислого газа стволом [13]. Например, при резком падении температуры воздуха на 5-10°С возникает значительный градиент между температурами ствола и окружающего воздуха, что, в свою очередь, может привести к резкому увеличению выделения СО2 из ствола [13]. По нашему предположению, наличие смоляных ходов в стволах хвойных может также сказаться на интенсивности дыхания сосны после пожара. Так резкое и значительное увеличение температуры в результате пожара может привести к плавлению смолы внутри смоляных ходов и их блокировке. Это приводит к задерживанию выделения углекислоты с поверхности ствола, и чем сильнее пирогенное воздействие, тем сильнее эффект.
В нашем исследовании дыхание ствола ослабленных деревьев после пожара увеличивается по сравнению со здоровыми и сильно ослабленными деревьями, но увеличивается в разной степени (рисунок 1). Так, на участке 3, где более низкая скорость распространения пожара реакция выше, чем на участке 5.
Анализ дыхания ствола до пожара показал (рисунок 1), что у здоровых деревьев по сравнению с ослабленными дыхание в 1.5 раза выше на 3-м участке (t=5.8, P=0.0002), и в 1.6 раза на 5-м участке (t=3.6, P=0.003). Достоверных различий в интенсивности дыхания между ослабленными и сильно ослабленными деревьями до выжигания не обнаружено (рисунок 1). Забуга [3] отметила сходную с нашей зависимость дыхания ствола сосны обыкновенной от жизненного состояния дерева в условиях лесостепной зоны Предбайкалья.
Так, ею показано, что в течение летней вегетации дыхание ствола сосны на высоте 1.3 м у деревьев II-го класса роста составило 6-10 мг СО2/(дм2 ч) (или 3.8-6.3 mмоль СО2/(м2 с)), у деревьев III-го класса роста – 4-6 мг СО2/(дм2 ч) (или 2.5-3.8 mмоль СО2/(м2 с)), у угнетенных – IV-го класса роста – 3-4 мг СО2/(дм2 ч) (или 1.9-2.5 mмоль СО2/(м2 с)).
Таблица 5. Интенсивность дыхания стволов сосны
№ дерева |
Жизненное состояние |
Температура воздуха до выжигания, оС |
Интенсивность дыхания до выжигания, µмоль СО2/(м2 с) |
Температура воздуха после выжигания, оС |
Интенсивность дыхания после выжигания, µмоль СО2/(м2 с) |
3-й участок |
336 |
I |
27.7±0.3 |
6.3±0.4 |
27.2±0.1 |
4.8±0.5 |
335 |
I |
28.1±0.3 |
7.6±0.5 |
29.6±0.0 |
5.6±0.3 |
320 |
II |
30.0±0.1 |
4.3±0.6 |
27.6±0.1 |
11.1±0.7 |
191 |
II |
28.5±0.0 |
4.6±0.1 |
28.0±0.1 |
8.4±0.5 |
326 |
III |
27.9±0.1 |
4.4±0.4 |
27.2±0.1 |
2.3±0.1 |
108 |
III |
28.2±0.1 |
4.6±0.3 |
29.1±0.2 |
4.5±0.2 |
Средняя по древостою интенсивность дыхания ствола, µмоль СО2/(м2 с) |
5.3±0.3 |
6.1±0.7 |
5-й участок |
161 |
I |
24.8±0.2 |
10.1±0.5 |
24.9±0.0 |
5.6±0.5 |
340 |
I |
25.3±0.0 |
6.4±0.8 |
27.0±0.1 |
5.8±0.4 |
312 |
II |
24.8±0.1 |
6.3±0.5 |
23.7±0.0 |
7.0±0.6 |
187 |
II |
24.5±0.1 |
3.7±0.7 |
22.6±0.0 |
6.2±0.2 |
150 |
II |
24.9±0.1 |
5.0±0.4 |
23.3±0.1 |
5.3±0.3 |
997 |
III |
25.1±0.0 |
5.4±0.5 |
25.5±0.1 |
3.2±0.3 |
302 |
III |
24.6±0.2 |
6.8±0.8 |
24.1±0.0 |
4.2±0.4 |
Средняя по древостою интенсивность дыхания ствола, µмоль СО2/(м2 с) |
6.3±0.4 |
5.3±0.3 |
Примечание: I - здоровое, II - ослабленное, III - сильно ослабленное. Приведены средние по интенсивности дыхания ствола для отдельных деревьев и по древостою и стандартная ошибка среднего.
Подобные данные встречаются в работах других исследователей [13], где дыхание ствола сосны в зависимости от жизненного состояния в течение летней вегетации у деревьев I-II-го класса роста варьирует от 0.25-0.45 мг СО2/(м2 с) (или 5.7-10.2 mмоль СО2/(м2 с)), у ослабленных (III-IV класс роста) – 0.08-0.38 мг СО2/(м2 с) (или 1.8-8.6 mмоль СО2/(м2 с)).
Многие исследователи обнаруживают увеличение интенсивности выделения СО2 с поверхности ствола с увеличением диаметра ствола [13]. В нашем случае, изучение дыхания ствола в зависимости от его диаметра показало, что на обоих участках внутри каждой выделенной группы жизненного состояния подобная связь отсутствует (рисунок 2).
Лишь в группе здоровых деревьев на 3-м участке дыхание ствола с увеличением диаметра несколько повышается (до 16%). В целом, на 3-участке дыхание прямо пропорционально возрастает с увеличением диаметра (R2=0.63), тогда как на 5-м участке такая связь отсутствует (R2=0.001). Наличие связи между диаметром ствола и его дыханием свидетельствует о том, что диаметр ствола в большинстве случаев является параметром, определяющим интенсивность дыхания в период активной вегетации деревьев.
Если пренебречь различиями в дыхании стволов сосны в зависимости от ее жизненного состояния, то их дыхание на рассматриваемых участках не отличалось между собой, как до выжигания (t=-1.8, P=0.08), так и после (t=1.2, P=0.25). Но при уровне значимости Р<0.1 (табл. 5) наблюдалось достоверное снижение интенсивности дыхания стволов на 5-м участке на 16% (t=2.0, P=0.06), на 3-м участке дыхание ствола не менялось после пожара (t=-1.1, P=0.29).
Это может означать, что среднее по древостою дыхание ствола при низовых пожарах низкой интенсивности не меняется или происходит его снижение.
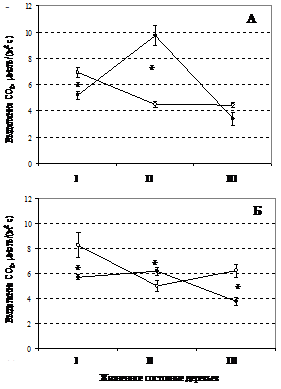 
Рисунок 1 - Интенсивность выделения углекислоты с поверхности ствола сосны на участке 3 (А) и 5 (Б). Приведены средние внутри группы жизненного состояния по данным табл. 5 и стандартная ошибка среднего. Примечание: * - различия достоверны при Р<0.05.
Данные исследования проведены для низовых пожаров низкой интенсивности, при пожарах же средней и высокой интенсивности будет наблюдаться иная реакция дыхания ствола, что требует проведения дополнительных исследований.
Выводы
Интенсивность дыхания стволов сосны обыкновенной в южнотаежных сосняках Нижнего Приангарья составила от 1.0 до 11.3 mмоль СО2/(м2 с). Анализ, проведенный на группах деревьев со сходной интенсивностью дыхания ствола, показал, что среднее дыхание ствола в группе здоровых деревьев в 1.5-1.6 раза превышало дыхание ослабленных и сильно ослабленных деревьев.
Различий в дыхании ствола между группами ослабленных и сильно ослабленных деревьев не обнаружено.
После пирогенного воздействия происходит снижение средней интенсивности дыхания ствола в группах здоровых и сильно ослабленных деревьев, в то время как в группе ослабленных деревьев дыхание возрастает. В целом, средняя интенсивность дыхания стволов в насаждении была сходной, как до выжигания, так и после.
 
Рисунок 2 - Интенсивность дыхания ствола в зависимости от диаметра дерева в различных группах жизненного состояния сосны на участках (А, Б) до выжигания. Приведены средние значения для деревьев и стандартная ошибка среднего.
На основании этих данных отмечены различия в дыхании ствола сосны до и после выжигания во всех взятых нами группах на обоих участках, за исключением сильноослабленных деревьев на 3-м участке (рисунок 1). У здоровых деревьев на обоих участках отмечается достоверное снижение дыхания ствола в 1.3 раза – на 3-м и в 1.4 - на 5-м участке.
У ослабленных деревьев на обоих участках наблюдается сходная реакция – достоверное усиление интенсивности дыхания на следующий день после пожара в 2 раза на 3-м участке и в 1.2 раза на 5-м участке. В то время как, у сильно ослабленных деревьев наблюдается тенденция к снижению дыхания ствола, которое на 3-м участке – недостоверно, а на 5-м участке дыхание ствола после выжигания достоверно ниже в 1.7 раза (рис. 1).
Колебания температуры в стволе связаны также с его объемом, объемом и составом газовой фазы в древесине, что может вызвать либо избыточное выделение, либо, наоборот, поглощение углекислого газа стволом [13]. Например, при резком падении температуры воздуха на 5-10°С возникает значительный градиент между температурами ствола и окружающего воздуха, что, в свою очередь, может привести к резкому увеличению выделения СО2 из ствола [13]. По нашему предположению, наличие смоляных ходов в стволах хвойных может также сказаться на интенсивности дыхания сосны после пожара. Так резкое и значительное увеличение температуры в результате пожара может привести к плавлению смолы внутри смоляных ходов и их блокировке. Это приводит к задерживанию выделения углекислоты с поверхности ствола, и чем сильнее пирогенное воздействие, тем сильнее эффект.
Библиографический список
- Utkin A.I., Zamplodchikov D.G., Korovin G.N., Chestnykh O.V. Reserves and density of organic carbon in forests of Russia // Proceedings of IBFRA 2000 Conference “The role of boreal forests and forestry in global carbon budget” / Eds Show C.H., Apps M.J. - Edmonton: Canadian Forest service, 2002. - P. 227-240.
- Lindroth A., Grelle A., Moren A.-S. Long term measurements of boreal forest carbon balance reveal large temperature sensitivity // Global Change Biology. - 1998. - № 4. - P. 443-450.
- Забуга Г.А. Эколого-физиологические аспекты продукционного процесса сосны обыкновенной в лесостепной зоне Предбайкалья: Дис. … докт. биол. наук. Иркутск, 2006. - 381 с.
- Lavigne M.B. Comparing stem respiration and growth of jack pine provenances from northern and southern locations // Tree Physiology. - 1996. - № 16. - P. 847-852.
- Waring R.H., Schlesinger W.H. Forest ecosystems concepts and management. New York: Academic Press Inc., 1985. - 340 p.
- Ryan M.G. The effect of climate change on plant respiration // Ecol. Appl. - 1991. - № 1. - P. 157-167.
- Ryan M.G., Waring R.H. Maintenance respiration and stand development in a subalpine lodgepole pine forest // Ecology. - 1992. - № 73. - P. 2100-2108.
- Davidenko E.P., Furyaev V.V., Sukhinin A.I., Goldammer J.G. Fire management needs in Russia’s boreal forest zone // The 3rd International Wildland Fire Congress, Sydney, Australia, 3-6 October 2003. http://www.wildlandfire03.com/home.asp.
- Иванова Г.А. Зонально-экологические особенности лесных пожаров в сосняках Средней Сибири: Дис. … докт. биол. наук. Красноярск, 2005. - 405 с.
- Классификация и диагностика почв России / Составители Шишов Л.Л., Тонконогов В.Д., Лебедева И.И. М.: Почвенный институт им. В.В. Докучаева РАСХН, 1997. - 235 с.
- Алексеев В.А. Диагностика жизненного состояния деревьев и древостоев // Лесоведение. - 1989. - № 4. - С. 51-57.
- Oohata S., Shidei T., Tsuji H., Hatakeyama I. Changes in respiratory rates of excised tree organs // Bull. Kyoto Univ. For. - 1967. - №39. - P. 100-109.
- Рост и газообмен СО2 у лесных растений / Цельникер Ю.Л., Малкина И.С., Ковалев А.Г. и др. М.: Наука, 1993. - 256 с.
- Ryan M.G. Growth and maintenance respiration in stems of Pinus contorta and Picea engelmannii // Can. J. For. Res. - 1990. - № 20. - P. 48-57.
- Shibistova O., Lloyd J., Zrazhevskaya G., Arneth A., Kolle O., Knohl A., Astrakhantceva N., Shijneva I., Schmerler J. Annual ecosystem respiration budget for a Pinus sylvestris stand in central Siberia // Tellus. - 2002. - № 54B. - P. 568-589.
- Sprugel D.G. Components of woody-tissue respiration in young Abies amabilis (Dougl.) Forbes trees // Trees. - 1990. - № 4. - P. 88-98.
- Han S.S., Suzaki T. Studies on the production of assimilates of trees. II. Seasonal changes of photosynthesis, respiration rates on the sun and shade leaves and estimation of branch replication by the living cell area method on the natural Japanese beech forest // J. Fac. Agr. Kushu Univ. - 1979. - V. 24. - № 2/3. - P. 133-144.
- Law B.E., Ryan M.G., Anthoni P.M. Seasonal and annual respiration of a ponderosa pine ecosystem // Global Change Biology. - 1999. - № 5. - P. 169-182.
- Stockfors J. Temperature variations and distribution of living cells within tree stems: implications for stem respiration modeling and scale-up // Tree Physiology. - 2000. - № 20. - P. 1057-1062.
- Malkina I.S., Yakshina A.M., Tsel’niker Yu.L. Relationship between CO2 evolution by the trunk and gas-exchange of the leaves in oak // Sov. Plant Physiology. - 1985. № 32. - P. 593-599.
- Lavigne M.B. Stem growth and respiration of young balsam fir trees in thinned and unthinned stands // Can. J. For. Res. - 1988. - № 18. - P.483-489.
- Martin T.A., Teskey R.O., Dougherty P.M. Movement of respiratory CO2 in stems of loblolly pine (Pinus taeda L.) seedlings // Tree Physiology. - 1994. - № 14. - P. 481-495.
- McGuire M.A., Teskey R.O. Estimating stem respiration in trees by a mass balance approach that accounts for internal and external fluxes of CO2 // Tree Physiology. - 2004. - № 24. - P. 571-578.
- Лакин Г.Ф. Биометрия. М: Высшая школа, 1990. - 350 с.
- Penning de Vries F.W.T. The cost of maintenance processes in plant cells // Ann. Bot. 1975. - № 39. - P. 77-92.
- Criddle R.S., Hopkin M.S., McArthur E.D., Hansen L.D. Plant distribution and the temperature coefficient of metabolism // Plant Cell Environ. - 1994. - № 17. - P. 233-243.
- Carey E.V., DeLucia E.H., Ball J.T. Stem maintenance and construction respiration in Pinus ponderosa grown in different concentrations of atmospheric CO2 // Tree Physiology. - 1996. - № 16. - P. 125-130.
- Wang W., Kitaoka S., Shi F., Sasa K., Koike T. Respiration rate of stems and roots of a larch plantation with special reference to the seasonal change in their cambium activity // Proceedings of the 9th symposium on the joint Siberian permafrost studies between Japan and Russia in 2000, Sapporo, 2001. - P. 42-49.
- McRae D.J., Conard S.G., Ivanova G.A., Suchinin A.I., Baker S.P., Samsonov Y.N., Blake T.W., Ivanov V.A., Churkina T.V., Hao W.M., Koutzenogij K.P., Kovaleva N.V. Variability of fire behavior, fire effects and emissions in Scotch Pine Forests of Central Siberia // Mitigation and Adaptation Strategies for Global Change. - 2006. - №1. - P. 45-74.
|


