|
"Хвойные бореальной зоны" 2004г. с.
38-47
ОСОБЕННОСТИ МЕТАБОЛИЗМА СОСНЫ ОБЫКНОВЕННОЙ В РАЗЛИЧНЫХ ЛЕСОРАСТИТЕЛЬНЫХ ЗОНАХ ЕНИСЕЙСКОГО МЕРИДИАНА
© И.Л. Милютина, Н.Е.Судачкова, Г.П. Семенова
Институт леса им. В.Н. Сукачева СО РАН, Красноярск, Россия
Работа выполнена при финансовой поддержке РФФИ (грант № 04-04-49418)
Определяли содержание крахмала, низкомолекулярных углеводов, общего и белкового азота и свободных аминокислот в образцах побегов, хвои, луба и камбиальной зоны ствола и скелетных корней 27-40-летних деревьев сосны обыкновенной Pinus sylvestris L. из разных частей ареала вдоль р. Енисей в интервале 53-65° с.ш. У сосны в наиболее оптимальных условиях южной тайги отмечены лучшие приросты в высоту и по диаметру, снижающиеся в средней тайге и лесостепи на 25-40%, в северной тайге - до 50%. В начале вегетационного периода у сосны из северной популяции повышена концентрация Сахаров и азотсодержащих веществ в камбии и белков практически во всех тканях, а также свободных аминокислот в проводящих тканях луба наряду с другими растущими тканями. Между популяциями сосны, произрастающей в разных лесорастительных зонах, обнаружены различия в составе, локализации и соотношении метаболитов и запасных веществ
The content of starch, low molecular carbohydrates, total and protein nitrogen and free amino acids in the samples of current shoots, needles, inner bark and cambial tissues of stems and rough roots of 27-40 year old trees of Pinus sylvestris L. from differenr area parts along Enisey river in an 52-65° N.lat. was determined. There are the best annual height and diameter increments of pine trees in optimal conditions of south taiga, they are lower by 25-40% in middle taiga and forest steppe and by 50% in north taiga. At the beginning of vegetation the north pine altered by the increased concentration of sugars and nitrogen containing compounds in cambium and protein content in all tissues, and increased amino acid concentration in conducting tissues of inner bark equally with other growing tissues. The differences in content, localisation and ratio of metabolites and reserve compounds among populations of pine from various forest zones were found
Введение
Негативное влияние природных факторов, лимитирующих рост и продуктивность хвойных в Сибири, во многом определяется действием водного и низкотемпературного стрессов. На протяжении ареала сосны обыкновенной в меридианальном направлении основными неблагоприятными факторами среды являются гипотермия, связанная с присутствием длительно-сезонной мерзлоты в зоне северной тайги и летние засухи в лесостепной зоне. Интенсивность процесса ксилогенеза зависит от обеспечения растущих древесных тканей основными метаболитами. Суровые условия существования вызывают существенные изменения в метаболизме растений, что приводит к изменению состава и соотношения метаболитов и запасных веществ и, как следствие, к изменению морфологических параметров деревьев [1,2].
В связи с этим задача исследования состояла в изучении особенностей метаболизма сосны
обыкновенной из различных лесорастительных зон вдоль Енисея.
Объекты и методы
Объектом исследования была сосна обыкновенная Pinus sylvestris L. в возрасте 27-40 лет из разных частей ареала вдоль р. Енисей. Подзона северной тайги представлена насаждениями в районе пос. Туруханск (65° с.ш.), в зоне средней тайги модельные деревья были отобраны в районе пос. Никулино (60° с.ш.), на южной границе таежной зоны объектами исследования были молодняки сосны вблизи г. Красноярска (56° с.ш.), в лесостепной зоне - насаждения в районе г. Минусинска (53° с.ш.). В начале вегетации в период активной камбиальной деятельности с 10 деревьев отбирали образцы хвои текущего года (хвоя I) и однолетней хвои (хвоя II), наружного (луб I)
и внутреннего слоя вторичного луба (луб II) стволов и скелетных корней, комплекса живых тканей обесхвоенных побегов текущего года (ксилему удаляли), камбиальной зоны ствола и корней. Наружный слой вторичного луба сосны представляет собой дилатационную зону,
насыщенную паренхимными элементами и смоловместилищами, внутренний проводящую часть. Камбиальная зона – это слой клеток, включающий как камбиальные инициали, так и растягивающиеся клетки вторичной ксилемы до начала лигнификации (отсутствие реакции с флороглюцином). Растительный материал фиксировали 80 % этанолом. В образцах проводили
определение общего азота колориметрически с реактивом Несслера, содержание свободных аминокислот определяли на аминокислотном анализаторе ААА-339. Крахмал экстрагировали хлорной кислотой и определяли иодометрически [3], концентрация низкомолекулярных углеводов определялась с использованием модифицированного метода Бертрана [4].
Результаты и обсуждение
При сравнении морфологических характеристик деревьев сосны из различных местообитаний наилучшие показатели отмечены у сосен, произрастающих вблизи Красноярска. У деревьев сосны в зоне средней тайги и в лесостепи прирост в высоту и по диаметру по сравнению с деревьями, растущими в оптимальных условиях, снижен на 25-40%, на северной границе ареала снижение этих показателей достигает двукратной величины (таблица 1).
Таблица 1 - Морфометрические характеристики модельных деревьев сосны обыкновенной
Параметр |
Лесостепь |
Южная тайга |
Средняя тайга |
Северная тайга |
Возраст, лет |
36 |
27 |
30 |
40 |
Высота, м |
9,2 ± 0,1 |
12,1 ± 0,5 |
8,4 ± 0,8 |
9,1 ± 0,2 |
Диаметр корневой шейки, см |
14,1 + 0,5 |
15,7 ± 0,7 |
11,2 ± 0,3 |
13,0 ± 0,2 |
Прирост в высоту, см |
26 |
45 |
28 |
23 |
Прирост по диаметру, мм |
3,9 |
5,2 |
3,4 |
2,9 |
Длина однолетнего побега, см |
12,2 ± 0,8 |
21,5 ± 0,7 |
12,5 ± 0,5 |
4,5 + 0,2 |
Длина однолетней хвои, см |
6,8 ± 0,1 |
7,0 ± 0,1 |
6,4 ± 0,1 |
3,5 ± 0,1 |
С продвижением с юга на север наблюдаются существенные изменения в составе растворимых углеводов, крахмала,общего и белкового азота и свободных аминокислот в тканях и органах сосны. В период активной камбиальной деятельности
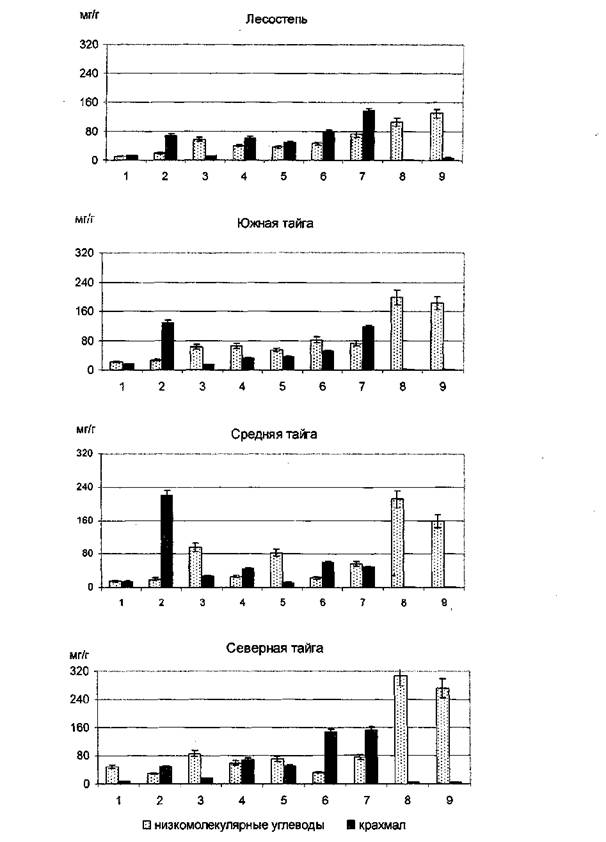
для обеспечения ростовых процессов необходимость в ассимилятах как углеводной, так и азотной природы очень высока. В это время наряду с синтезом соединений de novo идет и интенсивная мобилизация резервных веществ за счет перевода их в лабильные низкомолекулярные соединения. При этом следует отметить, что у деревьев, растущих в наиболее неблагоприятных климатических условиях, всегда отмечается достаточно высокая концентрация запасных веществ, в частности крахмала [5,6]. Наиболее существенная концентрация крахмала наблюдается в тканях луба корней и в однолетней хвое, хотя в разных лесорастительных районах крахмал по-разному перераспределяется между этими аккумулирующими зонами (рисунок 1). По содержанию растворимых лабильных Сахаров лидирующее положение занимают ткани камбиальной зоны ствола и корней сосны из северной части ареала, что может объясняться невостребованностью низкомолекулярных форм углеводов в процессах синтеза структурных элементов или нарушениями в транспорте. Концентрация общего азота в камбиальной зоне северной сосны снижена в 1,5-2 раза по сравнению с камбием сосен из других местообитаний (рисунок 2). В ассимилирующих органах следует отметить,
наряду с недостатком азотсодержащих соединений в однолетней хвое северного
варианта сосны, также пониженную их концентрацию в тканях побега у сосны, произрастающей в относительно оптимальных условиях. Снижение содержания белкового азота наблюдается практически во всех тканях и органах сосны северной популяции. Ранее было отмечено характерное для древостоев подзоны северной тайги меньшее количество азотсодержащих соединений как вследствие меньших запасов биомассы, так и пониженной концентрации их в органах и тканях [7].
Наглядным показателем стрессового состояния деревьев сосны может служить величина отношения неструктурных углеводов (суммы растворимых Сахаров и крахмала) к концентрации общего азота (таблица 2). В тканях луба ствола и корней сосны, растущей в суровых климатических условиях северотаежной зоны, этот показатель составляет 12-16, достигая в
камбиальной зоне 20, в то время как в более благоприятных условиях он, как правило, не
превышает 10. Обращает на себя внимание также достаточно высокая величина данного
показателя в запасающих тканях луба корней сосны в лесостепной зоне - 15,6, что может
свидетельствовать о создании резервов перед наступлением летнего засушливого периода.
Анализ отношения неструктурные углеводы: общий азот у двух видов лиственниц показал,
что у лиственницы Гмелина, произрастающей в районах с более жестким климатом, по сравнению с лиственницей сибирской его величина существенно выше и также может достигать 20 в отдельных тканях [8].
Повышенное содержание крахмала у северной сосны в этот период проявляется в очень высоком отношении крахмал: белковый азот в тканях луба корней - 19-22.
В метаболизме азота важная роль принадлежит аминокислотам. Общее содержание свободных аминокислот в различных тканях получали путем суммирования концентраций индивидуальных соединений. Наибольшая концентрация свободных аминокислот наблюдается в растущих тканях побега и (или) хвои текущего года и камбиальной зоны ствола, а у северных вариантов также в проводящих тканях луба ствола (рисунок 2). Если по суммарному содержанию аминокислот трудно выявить особенности, характерные как для отдельных тканей, так и комплекса природно-климатических условий места произрастания, то по составу этих соединений можно отметить существенные различия.
В камбиальной зоне ствола и корней деревьев сосны во всех местообитаниях доминирующее положение занимает у-аминомасляная кислота (ГАМК), составляя от 60 до 77% от общего пула аминокислот (таблицы 3-6). Для северного варианта сосны характерна существенно повышенная доля аспарагиновой кислоты в камбии ствола и корней и глутаминовой кислоты в камбиальной зоне ствола. Во всех популяциях, за исключением южнотаежной, увеличена доля аланина в стволовой части.
Хвойные бореальные зоны. 2004. Выпуск 2. |
Рисунок 1 - Содержание низкомолекулярных Сахаров и крахмала в тканях и органах сосны обыкновенной из разных лесорастительных зон в начале вегетационного периода: 1 - хвоя текущего года, 2 - хвоя прошлого года, 3 - побеги текущего года, 4 - луб I ствола, 5 - луб II ствола, 6 - луб I корней, 7 - луб II корней, 8 - камбиальная зона ствола, 9 - камбиальная зона корней
Таблица 3 - Содержание свободных аминокислот в различных морфологических структура* деревьев сосны обыкновенной из лесостепи в начале вегетационного периода, % от суммы
Аминокислота |
Хвоя |
Хвоя |
|
|
|
|
|
Камби- |
Камби- |
текущего года |
прошлого года |
Побег |
Луб I ствола |
Луб II ствола |
Луб I корня |
Луб II корня |
альная зона
ствола |
альная зона корня |
Цистеиновая |
1,1 |
4,1 |
0,5 |
2,5 |
5,3 |
1,8 |
7,6 |
0,7 |
0,6 |
Таурин |
0,1 |
0,9 |
0,2 |
0,6 |
0,8 |
0,4 |
0,6 |
0,2 |
0,1 |
Аспарагиновая |
4,9 |
16,2 |
2,4 |
9,4 |
4,8 |
9,7 |
4,5 |
3,6 |
0,8 |
Треонин |
3,0 |
0,6 |
1,2 |
0,5 |
0,4 |
0,9 |
0,3 |
1,6 |
1,6 |
Серии |
12,4 |
3,1 |
4,4 |
4,7 |
3,3 |
7,0 |
3,1 |
6,3 |
7,8 |
Аспарагин |
6,4 |
0,6 |
1,5 |
0,3 |
0,7 |
0,5 |
0,2 |
0,7 |
0,5 |
Глутаминовая |
3,7 |
14,3 |
3,2 |
17,5 |
8,4 |
21,3 |
16,3 |
1,0 |
1,0 |
Глутамин |
1,6 |
0,5 |
0,5 |
0,5 |
0,2 |
0,8 |
0,5 |
0,2 |
0,2 |
?-Аминоадипиновая |
0,1 |
0,3 |
0,1 |
0,1 |
0,1 |
0,3 |
0,0 |
0,1 |
0,1 |
Пролин |
3,0 |
3,6 |
з,з |
4,6 |
17,7 |
7,6 |
39,0 |
3,2 |
1,8 |
Глицин |
0,3 |
0,5 |
0,3 |
0,5 |
1,1 |
0,5 |
0,7 |
0,8 |
1Л |
Алании |
17,1 |
34,9 |
14,8 |
15,5 |
11,3 |
13,8 |
8,3 |
5,0 |
1,0 |
Цитруллин |
0,2 |
0,7 |
0,3 |
1,0 |
2,2 |
0,5 |
0,5 |
0,6 |
0,4 |
?-Аминомасляная |
0,3 |
0,3 |
0,2 |
0,4 |
0,3 |
0,2 |
0,2 |
0,3 |
0,2 |
Валин |
6,1 |
1,9 |
3,1 |
2,6 |
20,9 |
1,5 |
0,7 |
4,1 |
3,6 |
Цистин |
0,2 |
0,1 |
0,1 |
0,1 |
0,3 |
0,3 |
0,2 |
0,2 |
0,1 |
Цистатионин |
0,1 |
0,1 |
0,1 |
0,1 |
0,3 |
0,2 |
0,2 |
0,1 |
0,1 |
Метионин |
0,1 |
0,1 |
0,1 |
0,1 |
0,4 |
0,1 |
0,1 |
0,1 |
0,0 |
Изолейцин |
1,2 |
0,2 |
1,1 |
0,7 |
0,4 |
0,7 |
0,5 |
2,0 |
1,7 |
Лейцин |
1,1 |
0,3 |
1,4 |
0,5 |
0,5 |
0,8 |
0,5 |
2,2 |
2,5 |
Тирозин |
1,5 |
0,5 |
2,2 |
0,2 |
0,4 |
0,7 |
0,3 |
1,4 |
1,4 |
Фенилаланин |
1,2 |
0,6 |
1,7 |
0,2 |
0,4 |
0,7 |
о,з |
0,9 |
1,2 |
?-Аланин |
0,8 |
0,8 |
0,3 |
1,0 |
1,3 |
0,8 |
0,9 |
1,1 |
. 1,4 |
?-Аминомасляная |
0,1 |
0,3 |
0,1 |
0,5 |
0,2 |
0,2 |
0,2 |
0,5 |
0,5 |
?-Аминомасляная |
26,5 |
9,3 |
26,9 |
34,0 |
16,9 |
25,3 |
12,7 |
61,1 |
66,3 |
Орнитин |
0,1 |
0,6 |
0,2 |
0,5 |
0,5 |
0,5 |
0,5 |
0,3 |
0,3 |
Лизин |
1,2 |
0,7 |
0,7 |
0,5 |
0,3 |
0,4 |
0,2 |
0,8 |
1,1 |
Гистидин |
0,2 |
0,3 |
0,2 |
0,4 |
0,4 |
0,3 |
0,4 |
0,2 |
0,2 |
Аргинин |
5,3 |
3,5 |
29,1 |
0,3 |
0,2 |
2,6 |
0,6 |
0,6 |
2,5 |
Сумма аминокислот глутаминовой группы * |
40,4 |
32,5 |
63,4 |
58,4 |
46,0 |
58,6 |
70,1 |
67,1 |
72,4 |
Примечание *- сумма глутаминовой кислоты, глутамина, пролина, ?-аминомасляной кислоты, аргинина, орнитина и цитруллина
Снижение содержания белкового азота наблюдается практически во всех тканях и
органах сосны северной популяции, вследствие чего неиспользованный аминный азот фиксируется в составе непротеиногенной ГАМК.
Наибольшая концентрация свободных аминокислот наблюдается в растущих тканях побега и (или) хвои текущего года и камбиальной зоны ствола, а у северных вариантов - в проводящих тканях луба ствола.
Основная доля в общем пуле свободных аминокислот принадлежит дикарбоновым аминокислотам и связанному с ними аланину, отмечена специфичность в их содержании и соотношении в зависимости от ткани и места произрастания сосны.
Таблица 4 - Содержание свободных аминокислот в различных морфологических структурах деревьев сосны обыкновенной из южной тайги в начале вегетационного периода, % от суммы
Аминокислота |
Хвоя
текущего
года |
Хвоя
прошлого года |
Побег |
Луб I ствола |
Луб II ствола |
Луб I корня |
Луб II корня |
Камби-
альная
зона
ствола |
Камби-
альная
зона
корня |
Цистеиновая |
0,3 |
1,3 |
0,1 |
9,1 |
1,5 |
12,6 |
3,8 |
0,4 |
0,7 |
Таурин |
0,2 |
0,5 |
0,3 |
1,7 |
0,4 |
3,3 |
0,9 |
0,4 |
0,3 |
Аспарагиновая |
3,5 |
19,5 |
2,9 |
5,2 |
5,4 |
5,0 |
5,7 |
1,4 |
3,4 |
Треонин |
0,7 |
0,7 |
0,7 |
0,5 |
0,5 |
0,8 |
0,8 |
1,6 |
0,9 |
Серии |
5,1 |
2,9 |
2,8 |
3,4 |
4,5 |
3,2 |
5,0 |
3,4 |
4,0 |
Аспарагин |
3,4 |
0,9 |
3,0 |
0,3 |
0,4 |
0,6 |
0,5 |
0,8 |
0,5 |
Глутаминовая |
2,5 |
33,3 |
1,6 |
6,8 |
6,2 |
11,8 |
5,7 |
0,7 |
3,0 |
Глутамин |
1,1 |
0,4 |
0,4 |
0,4 |
0,3 |
1,2 |
0,3 |
0,2 |
0,1 |
?-Аминоадипиновая |
0,1 |
0,1 |
0,1 |
0,1 |
0,1 |
0,1 |
0,2 |
0,0 |
0,0 |
Пролин |
5,0 |
3,4 |
6,0 |
2,9 |
1,3 |
10,9 |
3,5 |
1,3 |
1,4 |
Глицин |
0,3 |
1,0 |
0,0 |
2,3 |
0,3 |
5,1 |
2,3 |
0,9 |
0,7 |
Алании |
17,5 |
20,6 |
12,6 |
16,0 |
12,3 |
7,9 |
7,6 |
0,7 |
2,5 |
Цитруллин |
0,6 |
0,5 |
1,3 |
0,8 |
0,4 |
0,8 |
0,4 |
0,4 |
0,1 |
?-Аминомасляная |
0,2 |
0,3 |
0,2 |
0,2 |
0,2 |
0,4 |
0,2 |
0,6 |
0,1 |
Валин |
2,8 |
1,8 |
1,7 |
0,9 |
0,9 |
0,6 |
0,9 |
2,8 |
1,7 |
Цистин |
0,1 |
0,1 |
0,0 |
0,1 |
0,1 |
0,3 |
0,1 |
0,1 |
0,0 |
Цистатионин |
0,0 |
00,1 |
0,0 |
0,1 |
0,1 |
0,2 |
0,1 |
0,0 |
0,1 |
Метионин |
0,1 |
0,1 |
0,0 |
0,0 |
0,0 |
0,1 |
0,1 |
0,0 |
0,0 |
Изолейцин |
0,5 |
0,3 |
0,5 |
0,2 |
о,з |
0,3 |
0,4 |
1,2 |
0,7 |
Лейцин |
0,7 |
0,4 |
0,9 |
0,3 |
0,4 |
0,4 |
0,6 |
2,3 |
1,4 |
Тирозин |
1,4 |
0,5 |
0,9 |
0,4 |
0,5 |
0,5 |
0,6 |
0,9 |
0,6 |
Фенилаланин |
1,1 |
0,7 |
0,1 |
0,5 |
0,6 |
0,7 |
0,5 |
0,8 |
0,6 |
?- Алании |
0,3 |
0,4 |
0,2 |
0,6 |
0,3 |
1,3 |
0,5 |
0,4 |
0,2 |
?-Аминомасляная |
0,1 |
0,2 |
0,1 |
0,3 |
0,1 |
0,5 |
0,2 |
0,3 |
0,1 |
?-Аминомасляная |
24,1 |
7,3 |
19,5 |
46,2 |
60,8 |
30,6 |
54,5 |
77,2 |
74,4 |
Орнитин |
0,2 |
0,6 |
0,3 |
0,2 |
0,3 |
0,4 |
0,5 |
0,1 |
0,7 |
Лизин |
1,2 |
0,6 |
0,8 |
0,2 |
0,3 |
0,3 |
0,3 |
0,7 |
0,6 |
Гистидин |
0,9 |
0,7 |
0,6 |
0,1 |
0,3 |
0,4 |
0,5 |
0,2 |
0,3 |
Аргинин |
25,7 |
1,0 |
42,5 |
0,3 |
1,1 |
0,8 |
3,7 |
0,7 |
0,8 |
Сумма аминокислот глутаминовой группы * |
59,1 |
46,5 |
71,6 |
57,6 |
70,4 |
56,5 |
68,8 |
80,6 |
80,5 |
Примечание *- сумма глутаминовои кислоты, глутамина, пролина, ?-аминомасляной кислоты,
аргинина, орнитина и цитруллина
Таблица 5 - Содержание свободных аминокислот в различных морфологических структурах деревьев сосны обыкновенной из средней тайги в начале вегетационного периода, % от суммы
Аминокислота |
Хвоя текущего года |
Хвоя прошлого года |
Побег |
Луб I ствола |
Луб II ствола |
Луб I корня |
Луб II корня |
Камбиальная зона ствола |
Камбиальная зона корня |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
Цистеиновая
Таурин
Аспарагиновая |
0,8
0,2 8,4 |
1,9 0,5 6,0 |
0,2
0,1
2,9 |
3,1 0,4 8,6 |
0,2
0,1
26,0 |
5,6 0,8
0,1 |
0,5
0,1
10,9 |
0,5 0,0 2,1 |
1,2 0,2 2,9 |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
Треонин |
2,4 |
1,2 |
1,0 |
1,0 |
1,0 |
0,2 |
1,2 |
1,2 |
2,0 |
Серии |
12,9 |
4,1 |
4,7 |
3,9 |
10,4 |
2,5 |
6,8 |
5,9 |
7,6 |
Аспарагин |
2,5 |
0,7 |
1,6 |
1,0 |
0,5 |
0,7 |
0,4 |
0,7 |
0,1 |
Глутаминовая |
14,0 |
23,0 |
8,1 |
28,0 |
18,2 |
31,0 |
29,9 |
1,3 |
1,6 |
Глутамин |
2,3 |
0,3 |
0,7 |
1,1 |
0,7 |
1,5 |
0,1 |
0,2 |
0,7 |
?-Аминоадипиновая |
0,1 |
0,1 |
0,0 |
0,1 |
0,0 |
0,1 |
0,0 |
0,0 |
0,0 |
Пролин |
5,6 |
4,1 |
3,9 |
7,4 |
0,5 |
24,9 |
1,2 |
0,7 |
0,9 |
Глицин |
1,7 |
1,3 |
0,4 |
0,7 |
0,4 |
0,5 |
0,4 |
1,0 |
1,9 |
Алании |
14,4 |
31,4 |
6,2 |
10,3 |
8,2 |
5,7 |
10,2 |
6,7 |
5,3 |
Цитруллин |
0,5 |
0,5 |
0,2 |
0,5 |
0,2 |
1,0 |
0,4 |
0,5 |
0,7 |
?-Аминомасляная |
0,2 |
0,5 |
0,2 |
0,2 |
ОД |
0,2 |
0,1 |
0,2 |
0,3 |
Валин |
3,8 |
1,3 |
0,9 |
1,0 |
2,0 |
0,5 |
1,7 |
2,4 |
2,5 |
Цистин |
0,1 |
0,1 |
0,1 |
0,3 |
0,0 |
0,1 |
0,1 |
0,1 |
0,0 |
Цистатионин |
0,2 |
0,1 |
0,1 |
0,2 |
0,1 |
0,2 |
0,0 |
0,1 |
0,0 |
Метионин |
0,1 |
0,0 |
0,0 |
0,1 |
0,0 |
0,0 |
0,0. |
0,0 |
0,0 |
Изолейцин |
1,0 |
0,4 |
0,2 |
0,5 |
0,6 |
0,2 |
0,5 |
1,1 |
0,8 |
Лейцин |
1,9 |
0,6 |
0,3 |
0,6 |
1,3 |
0,3 |
0,8 |
2,3 |
1,9 |
Тирозин |
2,1 |
1,3 |
0,4 |
0,3 |
0,5 |
0,1 |
0,3 |
0,9 |
0,9 |
Фенилшганин |
2,0 |
1,1 |
1,1 |
0,2 |
0,7 |
0,2 |
0,9 |
1,0 |
2,0 |
?-Аланин |
1,5 |
2,1 |
0,2 |
0,9 |
0,6 |
0,3 |
0,5 |
1,2 |
1,1 |
?-Аминомасляная |
0,5 |
0,3 |
0,1 |
0,4 |
0,1 |
0,1 ' |
0,1 |
0,2 |
0,1 |
?-Аминомасляная |
16,2 |
14,1 |
10,3 |
27,6 |
25,7 |
11,0 |
30,4 |
67,9 |
60,6 |
Орнитин |
0,5 |
0,5 |
0,3 |
0,3 |
0,6 |
0,1 |
0,2 |
0,1 |
0,3 |
Лизин |
1,8 |
0,9 |
0,1 |
0,6 |
0,7 |
0,4 |
0,6 |
0,9 |
1,4 |
Гистидин |
1,0 |
0,3 |
0,8 |
0,3 |
0,1 |
0,2 |
0,2 |
0,1 |
0,5 |
Аргинин |
1,4 |
1,2 |
53,9 |
0,6 |
0,5 |
0,3 |
0,4 |
0,6 |
1,3 |
Сумма аминокислот
глутаминовои группы * |
40,5 |
43,7 |
77,4 |
65,4 |
46,5 |
69,8 |
64,1 |
71,3 |
66,2 |
Примечание * - сумма глутаминовои кислоты, глутамина, пролина, ?-аминомаслянои кислоты, аргинина, орнитина и цитруллина
Таблица 6 - Содержание свободных аминокислот в различных морфологических структурах деревьев сосны обыкновенной из северной тайги в начале вегетационного периода, % от суммы
Аминокислота |
Хвоя
текущего года |
Хвоя
прошлого года |
Побег |
Луб I ствола |
Луб II
ствола |
Луб I корня |
Луб II корня |
Камби-
альная зона
ствола |
Камби-
альная
зона
Корня |
Цистеиновая |
0,1 |
0,6 |
0,3 |
1,6 |
0,9 |
8,1 |
8,8 |
1,5 |
0,2 |
Таурин |
0,0 |
0,2 |
0,2 |
0,4 |
0,3 |
0,9 |
2,8 |
0,1 |
0,2 |
Аспарагиновая |
2,2 |
5,5 |
4,7 |
6,2 |
14,1 |
8,5 |
6,6 |
7,8 |
6,9 |
Треонин |
0,9 |
1,0 |
1,5 |
0,3 |
0,7 |
0,8 |
0,2 |
1,0 |
0,7 |
Серии |
9,4 |
0,3 |
4,8 |
2,9 |
4,3 |
6,1 |
5,7 |
4,2 |
4,2 |
Аспарагин |
0,7 |
0,3 |
3,0 |
0,2 |
0,4 |
0,2 |
0,3 |
0,6 |
0,1 |
Глутаминовая |
8,9 |
31,4 |
10,3 |
48,5 |
33,6 |
36,6 |
26,0 |
5,2 |
2,3 |
Глутамин |
2,1 |
0,2 |
0,4 |
0,2 |
0,1 |
0,2 |
0,2 |
0,1 |
0,3 |
?-Аминоадипиновая |
0,1 |
0,1 |
0,2 |
0,1 |
0,1
___ - |
0,6 |
1,0 |
0,4 |
0,3 |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
Пролин |
2,4 |
2,6 |
6,8 |
3,4 |
4,7 |
11,3 |
12,3 |
1,7 |
1,1 |
Глицин |
0,7 |
1,2 |
1,1 |
1,5 |
1,5 |
2,3 |
1,4 |
0,8 |
0,7 |
Алании |
22,5 |
40,4 |
11,2 |
8,6 |
8,8 |
12,6 |
9,4 |
7,9 |
2,9 |
Цитруллин |
0,6 |
0,5 |
2,0 |
0,6 |
0,5 |
0,4 |
0,5 |
0,8 |
0,3 |
?-Аминомасляная |
0,3 |
0,2 |
0,3 |
0,3 |
0,2 |
0,3 |
0,3 |
0,7 |
0,3 |
Валин |
1,6 |
0,8 |
1,7 |
1,0 |
1,6 |
1,1 |
1,6 |
1,8 |
1,2 |
Цистин |
0,1 |
0,3 |
0,2 |
0,3 |
0,2 |
0,4 |
0,4 |
0,2 |
0,2 |
Цистатионин |
0,2 |
0,2 |
0,2 |
0,3 |
0,2 |
0,3 |
0,4 |
0,1 |
0,2 |
Метионин |
0,1 |
0,2 |
0,1 |
0,1 |
0,1 |
0,1 |
0,1 |
0,1 |
0,1 |
Изолейцин |
0,7 |
0,4 |
1,0 |
0,4 |
0,5 |
0,5 |
0,5 |
0,7 |
0,5 |
Лейцин |
0,8 |
0,5 |
1,0 |
0,4 |
0,4 |
0,6 |
0,6 |
1,5 |
0,8 |
Тирозин |
0,6 |
0,3 |
0,8 |
0,6 |
0,2 |
0,3 |
0,4 |
0,4 |
0,8 |
Фенил аланин |
0,5 |
0,4 |
0,8 |
0,6 |
0,2 |
0,3 |
0,3 |
0,4 |
0,9 |
?- Алании |
1,8 |
2,0 |
2,8 |
0,6 |
0,2 |
1,3 |
2,0 |
0,2 |
0,7 |
?-Аминомасляная |
1,0 |
1,2 |
1,8 |
0,4 |
0,1 |
1,2 |
1,8 |
0,1 |
0,6 |
?-Аминомасляная |
36,2 |
7,2 |
11,9 |
19,3 |
26,1 |
4,2 |
14,5 |
60,2 |
69,5 |
Орнитин |
0,3 |
0,4 |
0,8 |
о,з |
0,1 |
0,2 |
0,5 |
0,2 |
0,5 |
Лизин |
0,8 |
0,5 |
1,9 |
0,3 |
0,1 |
0,2 |
0,5 |
0,5 |
0,7 |
Гистидин |
0,2 |
0,3 |
0,5 |
0,2 |
0,2 |
0,2 |
0,3 |
0,3 |
0,4 |
Аргинин |
4,1 |
1,0 |
28,1 |
0,2 |
0,2 |
0,3 |
0,5 |
0,6 |
2,4 |
Сумма аминокислот
глутаминовои группы * |
54,7 |
43,1 |
60,3 |
72,6 |
65,4 |
53,1 |
54,5 |
68,8 |
76,4 |
Примечание - сумма глутаминовои кислоты, глутамина, пролина, ?-аминомасляной кислоты, аргинина, орнитина и цитруллина
Библиографический список
- Mohr H., Shopfer P. Plant Physiology.-Berlin Heidelberg: Springer-Verlag, 1995.- 629 p.
- Levitt J. Responses of plants to
environmental stresses. Vol. I. Chilling, freezing, and high temperature stresses. - N.Y., L: Acad. Press, 1980.- 497 p.
- Humphreys F.R., Kelly J. A method for determination of starch in wood // Anal. Chem. Acta.-1961.- V. 24.- № 1.- P. 66-70.
- Вознесенский В.Л., Горбачева Г.И., Штанько Т.П., Филиппова Л.А. Определение Сахаров по обесцвечиванию жидкости Фелинга // Физиол. растений.- 1962.- Т.9.- Вып.2.- С, 255-266.
- Судачкова Н.Е., Милютина И.Л., Семенова Г.П. Особенности депонирования и Использования резервных веществ северными популяциями сибирских видов хвойных // Сибирский экологический журнал. -2003.- Т.Ю.- № 6.- С. 721-726.
- Судачкова Н.Е., Романова Л.И., Милютина И. Л., Кожевникова Н.Н., Семенова Г.П. Влияние природных стрессов на уровень и распределение углеводов в тканях сосны обыкновенной в Сибири // Лесоведение.- 1994.- № 6.- С.3-9.
- Судачкова Н.Е., Милютина И.Л., Семенова Г.П., Плаксина И.В.Обеспеченность азотсодержащими соединениями основных лесообразующих хвойных пород. - В: Лесные экосистемы Енисейского меридиана.- Новосибирск. Изд-во СО РАН, 2002, -С. 311-318.
- Судачкова Н.Е., Милютина И.Л., Семенова Г.П. Специфика метаболизма лиственницы сибирской и лиственницы Гмелина в различных экологических условиях // Хвойные бореальной зоны. Вып. 1. Лиственница.- Красноярск, 2003.- С.54-60.
- Измайлов С.Ф. Азотный обмен в растениях. - М.: Наука, 1986.- 320с.
|