|
"Хвойные бореальной зоны" 2004г. с.
7-12 ИЗЪЯТИЕ ФИТОМАССЫ В НАСАЖДЕНИИ: ФЕНОМЕНОЛОГИЧЕСКАЯ МОДЕЛЬ
Исаев А.С.1, Суховольский В.Г.2, Бузыкин А.И.2, Овчинникова Т.М.2, Хлебопрос Р.Г.3
1Центр по проблемам экологии и продуктивности лесов РАН, Москва, Россия
2Институт леса им. В.Н.Сукачева СО РАН, Красноярск, Россия
3Институт биофизики СО РАН, Красноярск, Россия
Работа поддержана РФФИ (гранты 02-04-48769 и 04-04-49279)
Метод фазовых портретов используется для описания процессов роста фитомассы насаждения. На примере роста насаждения после проведения рубок ухода различной интенсивности показано, что процесс обладает марковскими свойствами и рост насаждения определяется текущим его состоянием и положением на фазовой плоскости.
The method of phase portraits is used for description of forest stand growth processes. On the example of stand growth after thinning cut of different volume it is shown that the process has Markov properties and the stand growth is determined by its current state and position on the phase plot.
Теоретические модели роста насаждений описывают изменения некоторой характеристики насаждения (фитомассы, числа деревьев и т.п.) во времени. Так как основным теоретическим положением является положение об ограниченности роста насаждения, то для описания роста насаждения используются уравнения ограниченного роста [5,7]. Обычно эти уравнения рассматриваются в интегральной форме как функция изменения фитомассы хво времени. Типичными примерами таких функций являются функция Ферлюльста (1) и функция Митчерлиха (2):
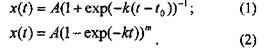
Из моделей (1) и (2) следуют следующие утверждения:
- достаточно, чтобы на без лесной территории появилось хотя бы одно дерево,
чтобы начался рост насаждения;
- насаждение, достигшее своей предельной фитомассы, будет находиться в этом состоянии неопределенно долго;
- при одномоментном изъятии части деревьев в насаждении характеристики рост
фитомассы насаждения не изменяется.
Первое из этих утверждений следует из факта неустойчивости точки х - 0; второе утверждение - из того факта, что А -устойчивая точка. Последнее утверждение следует из того, что в (1) и (2) при уменьшении значения у константы моделей не изменяются и движение продолжается по той же фазовой траектории.
Однако вытекающие из моделей (1) и (2) утверждения с точки зрения лесной экологии слишком категоричны. Во-первых, гибель насаждения - это столь же частый и естественный процесс, как и его рост, и процессы гибели должны описываться в модели. Во-вторых, неоднократно наблюдалось, что на территории занимавшейся ранее древостоями, которые затем полностью погибли после повреждения насекомыми-филлофагами, достаточно часто в течение длительного времени после гибели насаждений отсутствует возобновление не только хвойных, но и лиственных пород [13]. После повреждений сибирским шелкопрядом насаждений происходит сильное разрастание травянистых растений, задернение почвы, в результате чего гибнет весь уцелевший подрост и лесные площади превращаются в пустыри [6,8,12]. По данным В.В.Фуряева [13] на части участков шелкопрядников подроста и самосева хвойных практически нет, а лиственные возобновляются лишь отдельными куртинами. Размножению и росту травянистых растений, препятствующих развитию подроста, способствует резкое осветление территории, а состояние подроста тем хуже, чем лучше состояние травянистого покрова [14]. И, в- третьих, известно, что после рубок ухода, частичного повреждения деревьев насекомыми, слабых по интенсивности пожаров характер роста насаждения может существенно измениться по сравнению с тем, что наблюдалось до воздействия.
С помощью моделей (1) и (2) достаточносложно описать изменения процессов роста
насаждения под воздействием внешнихфакторов, таких, например, как климатические. Так, для (1) все внешние воздействия могут выражаться только как изменения параметров модели к и А. Но изменение удельной скорости роста к не будет приводить к изменению фазовой траектории, а лишь к изменению скорости движения системы по этой траектории. Только изменение предельной фитомассы А будет приводить к смещению фазовой траектории. При этом модель (1) не предсказывает каких-либо качественных изменений роста насаждений под воздействием внешних факторов.
Таким образом, модели типа (1) и (2) не позволяют описать многообразие процессов, происходящих в лесных насаждениях в ходе их роста и при различных внешних воздействиях на насаждения. В связи с этим необходим новый подход к описанию динамики роста лесных насаждений.
Для исследования процессов, связанных с устойчивостью лесных ценозов, реакцией
их на внешние воздействия, поиском стабильных или приводящих к разрушению состояний системы, оценкой критических величин изъятия фитомассы, приводящих к переходу из одного состояния системы в другое, удобнее использовать метод фазовых портретов, широко применяемый в физике, теоретической биологии и экологии [1,2]. Этот подход позволяет описать регуляторные механизмы формирования лесных биоценозов, запаздывание в регуляции и типы обратных связей [3].
На фазовой плоскости {х, х} можно не только отразить изменение состояния экосистемы, но и выделить на ней особые точки и области, при попадании в которые будет наблюдаться определенный тип поведения системы. В частности, такими особыми точками являются стационарные точки системы, то есть точки на фазовой плоскости, соответствующие состояниям системы, когда она не изменяется во времени. Если при отклонении значения х от стационарного значения х0 все изменения в системе направлены на возвращение х к значению хо к стационарному, то такая стационарная точка будет устойчивой [9].
Фазовый портрет системы характеризуется нуль - изоклинами, пересечения которых определяют его характерные точки. Выбор типа уравнений нуль-изоклин определяется особенностями изучаемой системы. Если возобновления леса на обезлесенной территории в обозримом будущем не происходит, то математически это означает, что нуль-изоклина лесной
экосистемы имеет локальный минимум и максимум с отрицательными и положительными значениями прироста фитомассы. Если процесс возобновления леса протекает успешно, то на фазовом портрете нуль - изоклина имеет локальный максимум, обусловленный взаимодействием деревьев в насаждении.
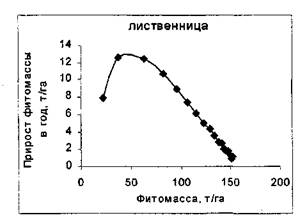
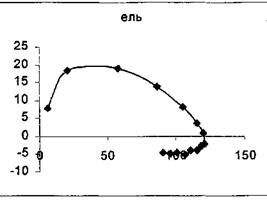
На рисунке 1 приведены примеры фазовых траекторий величин фитомассы лесов различных лесообразующих видов Северной Евразии, построенные по данным, приведенным в монографии В.А. Усольцева [11].
Лиственница (L. gmelinii), Среднесибирская Ель (P.obovata), Среднесибирская провинция,
провинция, Средняя тайга, возраст от 10 до 360 лет средняя тайга, возраст от 10 до 300 лет
Рисунок 1 - Фазовые портреты лесных насаждений
Для описания процессов роста насаждения с учетом конкуренции с травянистой растительностью ранее было введено следующее уравнение нуль-изоклин [3]:
х = kх * (А-х) * (х-В) + а
В (3) сомножитель (х-В) описывает процесс конкуренции древесной и травянистой растительности. Если фитомасса х древесных растений на территории превышает критическое значение В, то начинается рост насаждения. Если же (х - В) < О, то х < О и травянистые
растения вытесняют древесные.
Сомножители kx-(A-x) в (3), аналогичны уравнению (1) и описывают рост насаждения после преодоления конкуренции со стороны травянистых растений. Величина А в (3) как и в (1), характеризует максимальное значения фитомассы насаждения. Возобновление леса после его уничтожения может произойти только за счет занесенных семян, поэтому для описания процесса роста леса на безлесной территории в модель (3) добавлен член а > 0, характеризующий интенсивность роста фитомассы древесных растений, вырастающих на участке вследствие притока семян с других территорий.
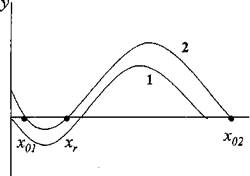
Фазовый портрет модели (3) роста насаждения, в которой учитываются эффекты конкуренции деревьев с травянистыми растениями и рост новых деревьев, приведен на рисунке 2 (кривая 2),
Рисунок 2 - Фазовые траектории уравнения роста (3). Кривая 1 - (а = 0); кривая 2 -{а = const > 0)
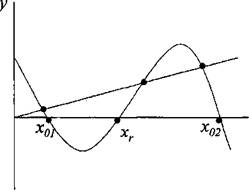
Рисунок 3 - Фазовый портрет насаждения с учетом постоянного изъятия или отпада части фитомассы
Видно, что при достаточно малых значениях а устойчивые состояния (3) соответствуют фитомассам х01 ? 0 и х02 = А, то есть можно говорить о двух устойчивых состояниях экосистемы на данной
территории. Первое из этих состояний соответствует экологической ситуации, когда деревья проигрывают в конкуренции с травянистыми растениями и территория превращается в луг, на котором появляются и гибнут древесные растения. Второе устойчивое состояние соответствует спелому насаждению, фитомасса которого достигает своего максимального значения. Для того чтобы на месте луга начал расти лес, согласно (3) необходимо, чтобы фитомасса деревьев превысила величину хг.
Кроме процессов роста, в любом насаждении идут процессы отпада и изъятия фитомассы. С учетом этого модель роста фитомассы можно записать следующим образом:
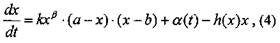
где значения функции изъятия h(x) находятся в интервале от 0 до 1 (при h(x)=0 изъятие отсутствует, при h(x) = 1 вся фитомасса насаждения при достижении определенного ее значения х отпадает или изымается).
В простейшем случае функцию изъятия можно записать как некоторую константу h
(0 < h < 1), предполагая, что в каждый момент времени изымается или отпадает определенная фиксированная доля фитомассы насаждения. В этом случае устойчивые состояния насаждения будут соответствовать точкам пересечения кривой х = kx • (А ~ х) • (х - В) + а и прямой х = hx(рисунок 3).
Как видно из рисунка 3, в зависимости от значения hвозможны два режима роста насаждения. Если будет выполняться
условие h < h0, то система будет иметь два устойчивых и одно неустойчивое состояние. При h> h0будет существовать только одно
устойчивое состояние, соответствующее ситуации, когда древесные растения поигрывают в конкуренции травянистым и роста насаждения нет. Таким образом, если по каким-либо причинам в насаждении увеличится отпад, это может привести к гибели древостоя.
Разного рода рубки, пожары, повреждения деревьев насекомыми также будут вести к изъятию части фитомассы. В простейшем случае разового изъятия фитомассы в некоторый момент времени то функция h(t) будет иметь следующий вид:

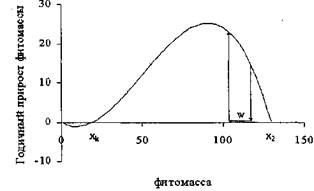
Такого рода изъятие производится в ходе промежуточных рубок, в частности, в ходе рубок ухода, когда одномоментно в древостое изымается часть фитомассы. При этом основной вопрос, интересующий лесоводов, состоит в том, насколько эффективны такие рубки и к какому возрасту принадлежит фитомасса насаждения. На рисунке 4 приведен фазовый портрет модели (16) с одномоментным изъятием фитомассы.
 Как видно из рисунка 4, рубки ухода привели к одномоментному уменьшению фитомассы на W единиц и точка, характеризующая текущее состояние насаждения, сместилась влево на фазовой траектории. Но после рубок ухода насаждение продолжало свое движение по той же фазовой траектории, что и до рубок ухода. Как видно из рисунка 4, рубки ухода привели к одномоментному уменьшению фитомассы на W единиц и точка, характеризующая текущее состояние насаждения, сместилась влево на фазовой траектории. Но после рубок ухода насаждение продолжало свое движение по той же фазовой траектории, что и до рубок ухода.
Рисунок 4 - Фазовый портрет модели (4) с одномоментным изъятием фитомассы
Таким образом, на фазовом портрете видно, что в модели (4) проведенные рубки
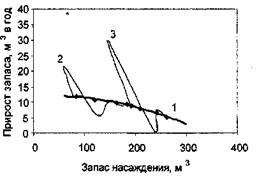
Рисунок 5 - Влияние рубок ухода на рост насаждений (1 - фазовая траектория не пройденного рубками ухода насаждения (контроль); 2 - фазовая траектория насаждения, у которого в возрасте 19 лет было изъято 22% фитомассы; 3 - фазовая траектория насаждения, у которого в возрасте 28 лет было изъято 20% фитомассы).
ухода не привели к качественным изменениям функции роста. Можно говорить, что уравнение (4) обладает марковскими свойствами и на поведение модели не влияет предыстория того, как модельный объект (насаждение) попало в соответствующую точку на фазовой траектории.
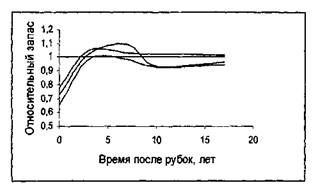
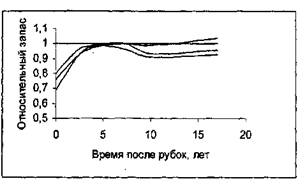
Представление о марковости процесса роста насаждения проверялась на данных о росте насаждений в течение 18 лет после рубок ухода [10]. На рисунке 5 приведены фазовые траектории насаждений после рубок ухода различной интенсивности (контрольные пробные площади объединены в один ряд; ход роста после рубок показан сплошными линиями для каждой пробной площади отдельно).
Как видно из рисунка 5, в конечном счете фазовая траектория насаждений после рубок ухода «выходит» на фазовую траекторию не пройденного рубками ухода контрольного насаждения, независимо от того, в каком возрасте производились рубки и какова была их интенсивность. Различие существует лишь во времени, необходимом для возвращения к фазовой траектории контрольного насаждений. Для насаждений, у которых интенсивность рубок ухода составила 20-22%, потребовалось 6-7 лет; насаждения с интенсивностью рубок 31-35% (рисунок 6).
А Б
Рисунок 6 - Динамика роста насаждений относительно контроля после рубок ухода. Возраст насаждения к моменту рубок: А -19 лет; Б - 28 лет
Из анализа модели (4) следует, что если разность между текущим значением фитомассы в момент времени и объемом изъятой фитомассы будет больше хг, то лес не погибнет и со временем компенсирует потерянную фитомассу, продолжая свой рост по соответствующей траектории на фазовом портрете. Если же после изъятия будет выполняться соотношение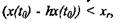 насаждение выйдет из области притяжения устойчивого состояния хог и на этой территории луг начнет преобладать над лесом. насаждение выйдет из области притяжения устойчивого состояния хог и на этой территории луг начнет преобладать над лесом.
Библиографический список
1. Арнольд В.И. Дополнительные главы теории обыкновенных дифференциальных уравнений,- М.: Наука, 1978.-304 с.
2. Исаев А.С.,Хлебопрос Р.Г. Принцип стабильности в динамике численности лесных насекомых//ДАН СССР.- 1973.- Т. 208.-№ 1.-С. 225-228.
3. Исаев А.С., Суховольский В.Г. Хлебопрос Р.Г., Бузыкин А.И., Овчинникова Т.М. Рост лесных насаждений: феноменологическая модель// Структурно- функциональная организация и динамика лесов. Красноярск.- 2004.- С. 309 - 311.
4. Исаев А.С., Хлебопрос Р.Г. Принцип стабильности в динамике численности лесных насекомых//ДАН.- 1973.- Т.208.- №1.- С. 225 - 228.
5. Карманова И.В. Математические методы изучения роста и продуктивности растений. - М.: Наука, 1976. - 222 с.
- Коломиец Н.Г. Сибирский шелкопряд и его роль в хвойных лесах Западной Сибири// Труды по лесному хозяйству Сибири. Новосибирск: Наука, 1962.-Вып. 7.-С. 137-161.
- Кофман Г.Б. Рост и форма деревьев.- Новосибирск: Наука, 1986. - 211 с.
- Кузьмичев В.В., Черкашин В.П., Корец М.А., Михайлова И.А. Формирование лесов на шелкопрядниках и вырубках в верховьях р. Большая Кеть (Красноярский край)// Лесоведение.- 2001.- № 4.- С. 8 - 14.
- Понтрягин А.С. Обыкновенные дифференциальные уравнения.- М.: Наука, 1970.-331 с.
- Рубцов В.И., Новосельцева А.И., Попов В.К., Рубцов В.В. Биологическая продуктивность сосны в лесной зоне.- М.: Наука, 1976.- 224 с.
- П.Усольцев В. А. Фитомасса лесов Северной Евразии: база данных и география.-Екатеринбург: УрО РАН, 2001. - 709 с.
- Фалалеев Э.Н. Естественное возобновление пихтовых лесов, поврежденных сибирским шелкопрядом//Лесной журнал.- 1960, № 2.-С. 35-37.
- Фуряев В.В. Роль пожаров в процессе лесообразования. Новосибирск: Наука, 1996.-253 с.
- Экологические аспекты лесовыращивания и лесопользования.-Новосибирск: Издательство СО РАН, 2001. -166 с.
|