|
Экологические системы динамичны по самой своей природе и постоянно испытывают на себе влияние изменчивости климата. Так как структура и функционирование лесных экосистем контролируются климатическими факторами через функции метаболизма, то в последующем могут измениться динамика восстановления лесов, прирост, ход роста и отпада деревьев, а также взаимоотношения пород. Ожидается, что изменение климата будет происходить быстрее, чем скорость, с которой экосистемы смогут адаптироваться и самовосстанавливаться.
Экосистемы Севера раньше всего должны среагировать на возможное глобальное потепление, поскольку климатическая обусловленность реакций экосистем проявляется здесь наиболее четко. В высоких широтах одновременно с ростом температуры воздуха должна увеличиться годичная норма осадков в связи с тем, что в теплые периоды пути прохождения циклонов сдвигаются к северу (Величко, 1992; Дроздов, Григорьева, 1971).
Следует ожидать значительных смещений границ распространения растительности. Считается, что скорость миграции различных видов деревьев в прошлом составляла порядка 4-200 км в столетие. В том, что касается регионов в средних широтах, то среднее потепление на 1-3,5 °С в следующие 100 лет будет означать перемещение существующих географических зон с одинаковыми температурами в направлении к полюсам примерно на 150-550 км или перемещение по высоте примерно на 150-550 м (Специальный доклад МГЭИК. Последствия изменения климата для регионов..., 1997).
В Финляндии в будущем столетии предполагаемое повышение температуры в высоких широтах на 2-4°C (следует рассматривать как фактор удлинения сезона роста), а также увеличение количества годовых осадков на 10-15 % позволит древесным породам расширить свои ареалы в направлении на север со скоростью 10-45 км за столетие (Year Book, 1996)
Глобальное потепление и уменьшение (или полное прекращение) пастьбы скота на альпийских лугах в Австрии привели к появлению многочисленных деревьев в полосе 100 м выше границы лесов (Stutzer, 1999). Многие из этих деревьев имеют повреждения в результате морозного засыхания и обледенения. В будущем следует ожидать подъема границы леса на 50 м, поскольку в этой полосе деревья отличаются удовлетворительным ростом и плодоносят. Однако климатические условия настолько суровы, что деревья в основном распространяются благодаря вегетативному размножению.
Соответственно состав видов растений в лесах, по всей вероятности, изменится. В некоторых регионах могут полностью исчезнуть целые типы лесов, и одновременно с этим могут возникнуть новые совокупности видов и соответственно новые экосистемы. На значительной части существующих лесных территорий земного шара (в среднем по земному шару на одной трети территорий с вариациями по регионам от одной седьмой до двух третей) в качестве одного из последствий возможных изменений в температуре и в наличии воды в условиях равновесия при эквиваленте удвоенного содержания CO2 произойдут крупные изменения в широко распространенных типах растительности; при этом наибольшие изменения произойдут в высоких широтах и наименьшие — в тропиках.
Прямое влияние на экосистемы будет оказывать увеличение содержания CO2 (а также ряда других газов) в атмосфере, которое может привести к повышению продуктивности и эффективности использования воды некоторыми видами растений. Вторичные воздействия изменения климата могут проявляться в изменениях характеристик почв и в экологических нарушениях (например: возникновение пожаров, распространение вредителей и болезней), которые будут более благоприятны для одних видов, чем для других, и приведут, в результате, к изменению состава и соотношения видов в экосистемах.
Варианты адаптации для экосистем ограничены, а их эффективность остается под вопросом. Соответствующие варианты содействия включают создание коридоров для содействия «миграции» экосистем, управление землепользованием, активное содействие естественному возобновлению, лесные культуры и искусственное восстановление деградировавших территорий.
Поскольку прогнозируемая скорость изменения климата будет больше скорости, с которой различные виды могут адаптироваться (в том числе и перемещение ареала), а также вследствие изоляции и фрагментации многих экосистем, наличия сопряженных факторов воздействия (природопользование, техногенное загрязнение, человеческая беспечность) и ограниченности вариантов адаптации, экосистемы (особенно лесные системы, горные системы и коралловые рифы) являются достаточно уязвимыми для воздействий изменения климата.
Как и в сценариях изменения климата, в прогнозах развития лесов встречаются как оптимистичные, так пессимистичные сценарии. Специальные исследования и модельные расчеты, проведенные в Институте глобального климата и экологии показали, что в ближайшие 50 лет влияние климатических изменений на рост лесов в целом будет небольшим - по порядку величины меньшим, чем влияние неравновесного возрастного распределения лесов, сложившегося под влиянием интенсивных рубок прошлых лет (Второе национальное сообщение..., 1998). С авторами можно согласиться только в том случае, если рассматривать исключительно прямое влияние изменения климата. Но в разрушении древостоев сочетание неблагоприятных климатических факторов, приводящее к ослаблению растений или созданию оптимальных условий для развития вредителей, болезней, пожаров лишь слабый сигнал. Ситуация даже в относительно благополучном 20 веке свидетельствует о огромной разрушающей роли вредителей, болезней, пожаров в бореальных лесах.
Результаты некоторых исследовании экосистем умеренной зоны Азии (между 18° с.ш. и Северным полярным кругом, включая: Японские острова. Корейский полуостров, Монголию, большую часть Китая и Российской Сибири), основанных на использовании различных моделей, дают основание считать, что в условиях климата с эквивалентно удвоенным содержанием CO2 произойдет значительное уменьшение площади (до 50 %) и продуктивности бореальных лесов (главным образом в Российской Федерации), сопровождаемое заметным расширением зон пастбищ и кустарников (Специальный доклад МГЭИК. Последствия изменения климата для регионов..., 1997). Произойдет также уменьшение площади зоны тундры (вплоть до 50 %), сопровождаемое выбросами метана из глубоких горизонтов торфяников и увеличением (до 25 %) выбросов CO2. Что также будет дополнительным фактором, способствующим увеличению парникового эффекта.
В случае потепления усиленный рост растений, при котором поглощается атмосферный диоксид углерода, не может компенсировать ускоренного разложения органических веществ. Так как при изменениях климата значительное потепление прогнозируется в северных и арктических районах, где огромные территории заняты тундрой и заболоченными землями. В свою очередь в зоне вечной мерзлоты при таянии льда все больше торфа будет подвергаться воздействию микроорганизмов, разлагающих органическое вещество и большое выделение СО2 и СН4 в атмосферу. По оценкам, при росте летней температуры в тундре на 4°С в атмосферу дополнительно выделится до 50% углерода из торфа, несмотря на более интенсивный рост растений это не предполагает компенсации (Кудеяров, 1994).
Некоторые заболоченные земли превратятся в леса или пустоши. Древесные растения, растущие на вечной мерзлоте, будут подвергнуты существенному воздействию в процессе ее таяния. Первоначальное общее воздействие потепления на накопление углерода в высокоширотных экосистемах, скорее всего, будет негативным, поскольку процессы распада вначале пойдут быстрее воспроизводства (Technical summary climate change 2001: A Report of Working Group II, 2001). В этих системах изменения альбедо и поглощения энергии в ходе зимы будут действовать как положительная подпитка регионального потепления в результате раннего таяния снега и, в ходе десятилетий и столетий, граница леса будет двигаться к полюсу. Большинство процессов в заболоченных областях зависят от гидрологических условий (дренажа); поэтому адаптация к предполагаемым изменениям климата может стать практически невозможной. Арктические и субарктические болотные сообщества на вечной мерзлоте, так же, как и более южные депрессионные заболоченные земли с небольшими дренажными участками, скорее всего, будут наиболее чувствительны к климатическим изменениям. В ряде случаев изменение дренажа может сопровождаться пожарами в торфяниках.
Районы вечной мерзлоты в настоящее время занимают 58% территории России. В Московском государственном университете им. М.В.Ломоносова был составлен прогноз изменения южной границы зоны мерзлоты на 2100 год. Огромные территории России в следующем веке будут находиться в районах деградации вечной мерзлоты. В южных районах Урала и Сибири границы многолетнемерзлых пород сдвинутся к 2100 году на 300-400 км в северном направлении.
В настоящее время тундровые почвы ведут себя следующим образом: во время периодов переувлажнения они - источник метана, во время засухи -сток. По поводу реакции тундры на изменение климата существуют различные прогнозы: так, если изменение климата приведет к уменьшению количества осадков в летний период, то тундра станет стоком метана, с другой стороны, имеются механизмы, стабилизирующие влажность почвы и температуру, которые компенсируют этот эффект. Таким образом, если изменение климата не стимулирует никакие сукцессионные изменения в тундровом ландшафте, то средний поток из тундры будет примерно таким же (Второе национальное сообщение..., 1998).
Исследования Государственного гидрологического института показывают, что потенциально заболоченные земли весьма уязвимы. Сейчас в результате процесса торфонакопления они являются нетто-стоком углерода. Он оценивается в, примерно, 150 млн.т С02-эквивалента в год. Даже небольшие антропогенные климатические воздействия на этот большой поток могут привести к существенным эмиссиям.
Исследованиями Института физики атмосферы РАН В краткосрочном временном масштабе поток метана из тундры может сильно реагировать на колебания климатических параметров, поскольку функционирование микроорганизмов сильно зависит и от температуры и от влажности почвы. В целом, в долгосрочной перспективе, в результате таяния вечной мерзлоты ожидается усиление эмиссии метана.
К настоящему времени нет однозначного мнения о направлении развития западносибирской природы. Большинство современных исследователей полагают, что идет прогрессирующая гидроморфизация (Романова, 1965; Нейштадт, 1971; Караваева, 1982; Константинов, Горожанкина, 1985). Однако существует иная точка зрения, согласно которой заболачивание в регионе к настоящему времени стабилизировалось и на некоторых участках сменяется разболачиванием (Орлов, 1975; Львов, 1976; Пьявченко, 1979; Глебов, 1988) Причина подобных разногласий кроется в том, что территориально-временная взаимосмена леса и болота представляют собой сложный и неоднонаправленный процесс, который определяется факторами и местного, и глобального порядка, меняющимися как в пространстве, так и во времени (Горожанкина, Константинов, 2000). Данное утверждение согласуется с нашими наблюдениями.
Деградация вечной мерзлоты западного бореального леса Канады обусловлена потеплением климата, при этом из-за повышения уровня воды лесные болота сменяются безлесными олиготрофными (Vitt et al., 2000). С момента малого ледникового периода деградировало лишь 9% территории вечной мерзлоты, поэтому большинство участков находится в реликтовом состоянии. Однако, состояние 22% сохранившейся вечной мерзлоты неустойчиво. Предполагается, что в существующих климатических условиях будет происходить дальнейшая утрата лесных территорий в бореальной континентальной зоне на западе страны, при этом прогнозируется продолжительное увеличение накопления органического вещества еще на 11%.
Влияние осадков на изменчивость прироста деревьев хвойных видов прослеживается во многих районах Субарктики. Однако оно более разнообразно, зависит от условий местообитаний и выражено в меньшей степени по сравнению с воздействием температуры воздуха. Часто это влияние опосредованно через почвенный гидрологический и термический режимы (Мазепа, 1999).
Не следует исключать возможность того, что увеличение зимних осадков, более поздний сход снежного покрова будет сопровождаться смещением на более поздние сроки активации камбия несмотря на увеличение летних температур (Ваганов, Кирдянов, Силкин, 1999).
Пространственное и временное распределение осадков и температур – главные факторы, определяющие распределение древесных растений на Земле. Температура воздуха влияет на физиологические процессы в растениях, и, в долговременном плане, на численность населения и уровень общественного развития. Температура определяет урожай и качество семян и фруктов, воздействуя на такие процессы, как цветение, распускание почек, созревание фруктов и шишек (Козловски, Палларди, 1997). Весенние и осенние изменения температуры воздуха также влияют на смягчение и отвердение хвои (Guak et al., 1998). Температура определяет процессы экосистемного уровня, такие как деградация и минерализация почвы. Непрямые эффекты, ассоциируемые с нагревом, могут быть даже более ощутимы, чем прямая связь температуры с ростом растений, - в субполярных биоценозах, где вероятен нагрев мерзлотного слоя (Mooney et al., 1999). Местные леса приспособлены к локальным особенностям климата. Однако, в случае значительного изменения климата, адаптация к местному неизменному климату может сыграть отрицательную роль для устойчивости.
Недостаток или избыток воды определяет течение наиболее важных процессов, связанных с биохимией питания. В частности, заболоченные леса чувствительны к изменениям гидрологического режима. Поскольку лесные экосистемы обычно занимают регионы, где нехватка влаги не имеет (или почти не имеет) место, распределение водных ресурсов в пространстве и времени представляется критическим для углеродного баланса и является одним из главных компонентов моделей, используемых для предсказания глобальных климатических изменений.
Изменения в распределении и численности растительных и животных видов отражаются в уровнях рождаемости и смертности, особенностях роста и распространения особей в популяции. Климат и почва определяют наличие и рост растений. Климат влияет на распределение и численность видов животных, определяя наличие ресурсов, плодовитость и выживание (Hansen, Rotella, 1999). Изменения в режимах воздействия, а также конкуренция и сотрудничество с другими видами также влияют на распределение животных и растений. Исключительно важную роль также играет человеческая деятельность.
Глобальное изменение климата может, прежде всего, повлечь изменение взаимоотношений между породами. Вероятнее всего, эти перемены проявятся на стадии лесовозобновления в зоне таежных лесов. На северной границе лесов лиственница является первопроходцем, но высокая требовательность к условиям освещения не дает ей возможности конкурировать с другими породами в тех районах, где для них имеются приемлемые климатические и почвенные условия. Поэтому при изменении климата лиственница на части своего ареала может смениться елью и пихтой, а в ряде случаев и сосной (Второе национальное сообщение, 1998). Для ели глобальное повышение температуры само по себе не сможет изменить условий роста и воспроизводства. Однако, изменение количества осадков может оказать на нее негативное влияние, особенно в зонах южной тайги и смешанных лесов. Для лиственных пород вероятное изменение климата будет менее значимо, чем для хвойных – по непосредственному воздействию. Однако, уничтожение хвойных, стимулируемое климатическими изменениями (вспышки массового размножения вредителей и болезней, пожары и др.), освобождает пространство для развития осины, березы, ольхи, ивы (косвенное воздействие).
При изучении влияние изменения климатических условий на леса и естественные охраняемые территории в Мексике отмечается ухудшение условия для существования растительности (повышение аридности) (Villers-Ruiz, Trejo-Vazquez, 1998). Ожидается исчезновение 13% лесов умеренно-холодного и умеренно-жаркого климата. С другой стороны, увеличится доля сухих тропических лесов, а также тропических лесов гумидных и субгумидных.
Вертикальное продвижение пихты субальпийской и ели Энгельмана отмечается на хорошо прогреваемых горных склонах Нью-Мексико (Dyer, Moffett, 1999). Оценка произведена с использованием серии аэрофотоснимков с 1930-х гг. На северо-восточном склоне горы граница леса и луга относительно стабильна.
При моделировании климатических преобразований во Французских Альпах и Французском Средиземноморье (повышение средней температуры на 3°C и незначительное увеличение объема осадков) на радиальный рост 5 древесных пород в 24 популяциях установлено, что только немногие из 24 популяций оказались чувствительны к изменениям климата (Keller et al., 2000). Все они расположены на границе их экологического ареала и могут быть разделены на две группы. Первая состоит из высокогорных популяций и реагирует увеличением роста, индуцируемым потеплением климата во время вегетационного периода. Вторая состоит из одной популяции сосны обыкновенной в Средиземноморье и реагирует сильным снижением роста из-за летнего водного стресса. Однако, индивидуальная реакция деревьев в виде радиального прироста без учета ценотических и экосистемных связей не может хорошо прогнозировать динамику популяции. Особенно это касается популяций, вошедших в первую группу.
При изучении реакции растений на изменение климата чрезвычайно важным является фенологическая реакция. В литературе отмечается отсутствие однозначной ожидаемой реакции на глобальное потепление (Минин, Гутников, 2000).
До определенного предела биота демпфирует направленные воздействия внешних факторов (в нашем случае климата) без существенных изменений структуры и целостности, и внешне ее реакция может достаточно продолжительное время в явной форме не проявляться или даже возможна обратная реакция. Очевидно, что на протяжении эволюции многочисленные вариации климата в ту или другую сторону переживались биотой именно таким способом (Ведюшкин, Колосов, Минин, Хлебопрос, 1995; Ведюшкин, Колосов, Минин, Хлебопрос, 1995).
В связи с потеплением климата, продолжающимся на Аляске с начала 1800-х гг., северная граница леса, образуемая елью, продвинулась в более высокие широты (Juday et al., 1999). Распространение деревьев дальше на север или выше в горы сдерживается лишь недостаточной приживаемостью самосева, чему препятствует и тундровый покров прилегающих к лесной границе территорий. Потепление климата в сочетании с увеличением количества летних осадков способствует продвижению ели в некоторых районах. Однако там, где осадков выпадает недостаточно (граница леса, проходящая по небольшим возвышенностям рядом с лугопастбищными угодьями) можно наблюдать исчезновение ели и наступление лугов или осинников.
Следует иметь ввиду установленную в географических культурах (Корткерос, северная Россия) закономерность - развитие семян в северных популяциях сосны обыкновенной и ели европейской происходило несколько быстрее, чем в южных популяциях (Fedorkov, 2001). Согласно модели Сарваса деревья, переносимые из более холодных регионов с более коротким сезоном роста в более теплые регионы с более продолжительным сезоном роста, развивают флоральные органы раньше, чем переносимые из более теплых регионов или местные породы.
В целом можно выделить ряд параметром ответной реакции лесных экосистем на изменение климата:
- Смещение зон в т.ч. в горах;
- Потеря лесов сопровождается определенным ущербом для гидрологических систем, увеличением эрозии почв;
- Разрушение лесов, вызывающее выброс углерода в атмосферу, может способствовать глобальному потеплению;
- Уменьшение биоразнообразия;
- Изменение продуктивности;
- Увеличение экстремальности увлажнения (заболачивание, иссушение);
- Рост пожарной опасности;
- Снижение количества и качества семян;
- Влияние на возобновление (особенно хвойных);
- Увеличение вредоносности болезней и вредителей;
- Снижение устойчивости из-за увеличения частоты неблагоприятных краткосрочных явлений (периодов аномально теплой погоды и заморозков, сильных ветров, снегопадов и т.п.);
Хотя позитивная реакция деревьев на возрастание концентрации углекислоты часто преувеличивается – если не принимаются во внимание абиотические и биотические экологические факторы, большинство экосистем, тем не менее, реагируют положительно в смысле повышения выхода углерода в ответ на увеличение углекислоты в атмосфере по сравнению с текущим уровнем окружающей среды. Увеличение концентрации углекислоты в теплицах служит известным приемом повышения интенсивности и продуктивности фотосинтеза.
Удвоение концентрации CO2 в атмосфере может непосредственно влиять на деревья через: 1) снижение обмена через устьичный аппарат и 2) усиление фотосинтеза. В большинстве случаев это приведет к усилению прироста деревьев и эффективности водопоглощения.
Исходя из физиологических особенностей пользу от повышения СО2 могут извлечь С3-растения, к которым относятся практически все деревья и многие из основных сельскохозяйственных культур: рис, пшеница, картофель, бобовые.
У С3-растений на первой стадии фиксации молекула СО2 связывается с рибулозодифосфатом, содержащим 5-углеродный сахар. В результате реакции, происходящей под действием фермента рибулозодифосфаткарбоксилазы, образуется короткоживущее нестабильное соединение, включающее 6-углеродный сахар. Оно распадается на два производных, которые содержат по три атома углерода - отсюда и название “С3-растения”. С диоксидом углерода за активный центр рибулозодифосфаткарбоксилазы конкурирует кислород атмосферного воздуха. Если побеждает О2, растение теряет энергию, так как во время утилизации кислорода не происходит фиксации СО2. По мере же увеличения концентрации углекислого газа вероятность его “выигрыша” в конкуренции с О2 за связывание с активным центром фермента повышается. Действительно, в ряде экспериментов, когда концентрация СО2 устанавливалась на уровне 600 ppm, фотореспирация снижалась на 50%, а ее ограничение означает, что растение может использовать больше своей энергии на построение тканей. Однако у этих растений в условиях возросшей концентрации СО2 повышенный фотосинтез наблюдается в начальной стадии экспериментов, но после временной активации наступает его торможение. Транспортная система растения полигенна, зависит от многих факторов (энергетических, гормональных и др.) и не может быстро перестроиться. Поэтому при длительном воздействии на растение СО2 в условиях повышенной концентрации фотосинтез снижается из-за избыточного накопления крахмала в хлоропластах (Мокроносов, Гавриленко, 1992).
Высокое содержание СО2 в воздухе может быть неблагоприятным для так называемых С4-растений, первые продукты фотосинтеза которых - соединения из четырех атомов углерода: яблочная и аспарагиновая кислоты, оксалоацетат. К этому классу относятся многие травы сухих, жарких тропических и субтропических областей, сельскохозяйственные культуры - кукуруза, сорго, сахарный тростник и др. У С4-растений имеется добавочный механизм карбоксилирования - своеобразный насос, концентрирующий СО2 вблизи активного центра фермента, позволяющий этим растениям хорошо расти при обычных концентрациях диоксида углерода. У С4-растений в обычных условиях энергозатраты на фотореспирацию значительно ниже и эффективность фотосинтеза поэтому выше, чем у С3-растений. Примерно то же происходит и при фотосинтезе, характерном для типичных суккулентов. Его называют САМ-фотосинтезом (Crassulacean Acid Metabolism). САМ-растения подобно С4-растениям используют и С3, и С4-пути фотосинтеза, но отличаются от С4-растений тем, что для них характерно разделение этих путей только во времени, но не в пространстве, как у С4-растений.
Отмечаемое в литературе снижение интенсивности фотосинтеза после его первоначального увеличения до уровня, что наблюдается у растений, живущих в атмосфере с нормальным газовым составом, находит объяснение в регуляторном действии углекислого газа на ростовую функцию растения (Алексеев, Киселева, Чернова, 1999). Длительное выдерживание растения при высокой концентрации СО2 сопровождается увеличением площади листьев, стимуляцией роста побегов второго порядка, относительным возрастанием доли корней и запасающих органов в растении, усилением клубнеобразования. Ростовая функция усиливается за счет формирования нового фотосинтетического аппарата. Это свидетельствует о “двойной” роли СО2 как субстрата в процессе фотосинтеза и как регулятора ростовых процессов. При повышении уровня углекислого газа в атмосфере устанавливается новое стационарное состояние системы, соответствующее новому уровню углекислоты, что и приводит к росту урожая преимущественно за счет увеличения объема всей фотосинтетической системы и в меньшей степени за счет интенсивности фотосинтеза на единицу площади листа.
Поскольку атмосферный углекислый газ частично поглощается осадками и поверхностными пресными водами, в почвенном растворе повышается содержание СО2 и как следствие этого происходит подкисление среды. При культивировании проростков пшеницы 2-3-кратное превышение над нормальным содержания растворенного углекислого газа сопровождалось торможением роста корней растений с изменением их морфологии (Алексеев, Киселева, Чернова, 1999). Первоначальное повышение концентрации СО2 в питательной среде вело к стимулированию наземной и корневой массы растений пшеницы.
Таким образом, с увеличением концентрации углекислоты С3-растения оказываются в более выгодном положении, чем С4- и САМ-растения, а это в свою очередь может иметь весьма серьезные последствия. Многие С4-растения станут редкими, или им грозит вымирание. В агроэкосистемах при выращивании С4-растений, например кукурузы или сахарного тростника, повышенная концентрация СО2 может привести к падению их продуктивности, преимущество же получат сорняки, которые представлены в основном С3-растениями (Алексеев, Киселева, Чернова, 1999). В результате возможно значительное снижение урожая.
Что касается прогнозов урожайности основных сельскохозяйственных культур для России, то происходящий рост средних приземных температур воздуха и рост СО2 в атмосфере, казалось бы, должны иметь положительный эффект. Воздействие только роста углекислого газа в атмосфере может обеспечить рост продуктивности ведущих сельскохозяйственных культур - С3-растений (хлебных злаков, картофеля, свеклы и др.) - в среднем на 20-30% , тогда как для С4-растений (кукурузы, проса, сорго, амаранта) этот рост незначителен (Мокроносов, 1994).
Многофакторность взаимодействия в биосфере затрудняет моделирование процессов и не позволяет однозначно говорить о положительном эффекте роста СО2. Было установлено, что насыщение приземного слоя атмосферы толщиной до 1 м избытком СО2 может откликнуться “эффектом пустыни”. Этот слой поглощает восходящие тепловые потоки, поэтому в результате его обогащения диоксидом углерода (в 1.5 раза в сравнении с нынешней нормой) локальная температура воздуха непосредственно у земной поверхности станет на несколько градусов выше средней температуры. Интенсивность испарения влаги из почвы увеличится, что приведет к ее иссушению (Скурлатов, 1994).
Эксперимент, в котором исследовалось воздействие роста содержания углекислого газа в атмосфере на деревья, выявил существенные изменения в физиологических процессах, росте и накоплении биомассы (Mooney et al., 1999; Aber et al., 2001). При изменении концентрации СО2 в широких пределах, в растениях, представляющих наиболее северные леса умеренной полосы, усилился фотосинтез (Eamus and Jarvis, 1989; Bazzaz, 1990; Mohren et al., 1996; Long et al., 1996; Kozlowski and Pallardy, 1997). Обзор экстенсивных экспериментов по изучению углекислотного обогащения показала различную, но позитивную реакцию в процессе накопления растительной биомассы. (Ceulemans and Mousseau, 1994; Saxe et al., 1998; Mooney et al., 1999).
В экспериментах, не предусматривавших экологический стресс, накопление биомассы было существенней для хвойных (130%), чем для лиственных видов (только 49%-й рост) при повышенном содержании СО2 (Saxe et al.,1998). Широкий спектр реакций растительности включал, в частности, взаимодействие отклика на повышение концентрации углекислоты с другими экологическими факторами (Johnson et al., 1998; Curtis and Wang, 1998; Mooney et al., 1999; Stitt and Krapp, 1999; Morison and Lawlor, 1999). В последних полевых экспериментах получено существенное (25%) ускорение роста леса при постоянно увеличивающейся концентрации СО2 для Pinus taeda L. в Северной Каролине (Delucia et al., 1999).
Важный вопрос, - как долговременны такого рода усиленные реакции? Наблюдавшаяся акклиматизация или понижающая регуляция уровней фотосинтеза (Long et al., 1996; Lambers et al., 1998; Rey and Jarvis, 1998) были отнесены к физиологическому отклику - накопление фотосинтетических резервов (Bazzaz, 1990) или морфологическому отклику - изменения у деревьев (Pritchard et al., 1998; Tjoelker et al., 1998). Снижение уровней фотосинтеза объяснялось также как результат водного стресса или стресса питания, возникающего в выращенных в горшках саженцах в связи с ограничением роста корней (Will and Teskey, 1997a; Curtis and Wang, 1998). Понижающая регуляция была зафиксирована в низкотемпературных системах (Арктическая тундра), где изначальный рост общего выхода углеродной массы снижаются после трех лет пребывания в атмосфере, обогащенной углекислотой (Shaver et al., 1992; Mooney et al., 1999). Понижающая регуляция вероятна в регионах, где доступ продуктов питания не усиливается с ростом содержания углекислого газа. Последние обобщения данных крупномасштабных полевых экспериментов, связанных с воздействием углекислоты показали, что, с учетом некоторых отклонений, реакция деревьев на СО2 – для этих краткопериодических исследований – постоянна (Norby et al., 1999). Была исследована также реакция деревьев близ поверхностных выходов глубоких геотермальных струй; установлено, что 26-процентный рост сохранялся в течение трех десятилетий воздействия атмосферы с повышенным содержанием углекислоты.
В краткосрочном плане, изменения в физиологии растений, похоже, являются основной реакцией на повышенное содержание углекислоты, сильно зависящей от местных природных условий, таких, как водный стресс, наличие питательных элементов, индивидуальными видовыми особенностями (Egli and Korner, 1997; Tjoelker et al., 1998; Berntson and Bazzaz, 1998; Crookshanks et al., 1998; Kerstiens, 1998). В долгосрочной перспективе, видовое разнообразие лесов определяет конкретную реакцию последних (Mooney et al., 1999).
В условиях повышенного содержания углекислого газа, эффективность использования воды (WUE) нарастает, что приводит к росту влажности почвы. Повышенная влажность почвы – как показано – является важным фактором повышения выхода углеродной массы в водно-лимитированных экосистемах (Mooney et al., 1999). Хотя еще нет ясности в вопросе о том, уменьшается ли устьичная проводимость при росте концентрации СО2, (Long et al., 1996; Will and Teskey, 1997; Saxe et al., 1998; Curtis and Wang, 1998), WUE нарастает вне зависимости от устьичной проводимости (Aber et al., 2001). С постоянной проводимостью, возросшая концентрация атмосферной углекислоты приводит к ускорению накопления углеродной массы при постоянной потере воды. Если проводимость уменьшается, складывается баланс между нарастанием накопления углерода (которое несколько замедляется за счет падающей проводимости) и снижением потери воды (также замедляемой за счет уменьшенной проводимости). Экспериментальные исследования особо фокусировались на реакции на уровне листьев. Надо сказать, что физиологическая реакция, наблюдаемая на одном уровне (лист), не обязательно предполагает аналогичную реакцию на уровне более высоком (полог, водосборный бассейн). Имеются свидетельства того, что сокращение устьичной проводимости у древесных саженцев не имеет места у взрослых деревьев (Ellsworth, 1999; Mooney et al., 1999; Norby et al., 1999).
В условиях повышенной температуры воздуха и удвоенной концентрации углекислоты отмечается усиленная гомеостатическая приспособленность лиственных пород деревьев по сравнению с таковыми хвойных (Максимов, Койке, 1999). Продукционный процесс якутских и японских популяций деревьев в условиях возможного потепления климата будет в основном лимитирован эндогенными факторами - устьичной проводимостью, а также экзогенными - обеспеченностью растений влагой и минеральными органогенами, особенно азотом.
Наиболее спорными представляются данные о благотворности роста СО2 для древесных растений. Повышение концентрации СО2 в атмосфере даже без учета глобального потепления способно привести к значительному изменению структуры и функционирования экосистем, что может быть неблагоприятно для растений. Положительная реакция на повышенное содержание углекислого газа в воздухе, наблюдаемая у отдельного растения, вовсе не обязательно означает, что будет иметь место усиленный рост растительных сообществ в целом.
Изменение аллокации углерода в растениях ведет к серии структурных и функциональных изменений (водный режим, транспирация, осморегуляция), которые определяют засухоустойчивость древесных пород (Guehl et al., 1999). Таким образом, эти явления должны быть учтены при прогнозировании реакций древесных пород на изменение климата, в частности, на более засушливые летние периоды в зоне умеренного климата.
Изменение концентрации СО2 должно быть строго сбалансировано с потреблением азота, фосфора, других питательных веществ, света, воды в продукционном процессе без нарушения экологического равновесия (Алексеев, Киселева, Чернова, 1999). Нарушение баланса безусловно скажется на устойчивости и продуктивности.
Многолетние изменения прироста по площади сечения, сопоставленные с динамикой температуры северного полушария, выявляют региональные особенности этой связи: в субарктической зоне прирост по площади сечения тесно связан с изменениями температуры северного полушария (R= +0.81-0.93), в подзоне северной тайги , связь между многолетними изменениями прироста по площади сечения и температурой отрицательная (R= -0.83-0.92) за исключением наиболее теплого периода (1925-1965 гг.), для которого связь положительная (R = +0.80-0.87), в подзоне средней тайги связь практически отсутствует, но в период потепления так же положительная (R= +0.63-0.71) (Шашкин, Ваганов, 2000).
Реконструкция годичных температур в северном полушарии на основе ширины годичных колец деревьев Северной Америки, Скандинавии, Сибири и Монголии показывает, что годичные температуры возрастали в начале XX века, достигнув пика к середине 1940-х гг., затем они несколько понизились в 1950-е и 1960-е гг., но в 1990-е гг. достигли максимального уровня за последние три века (Juday, Jacoby, D'Arrigo, 1999). В настоящее время стресс из-за засухи, отчасти возникшей благодаря повышению температуры, является фактором, ограничивающим рост деревьев во многих северных местообитаниях. Нынешнее потепление в сочетании с более засушливыми сезонами в некоторых местообитаниях изменяет реакцию части бореальных лесов на климат. Если в прежние столетия самые северные деревья бореального леса увеличивали прирост в ответ на потепление климата, то теперь некоторые из них лучше растут в более прохладные годы, чем в более теплые.
Для систематизации многочисленных результатов исследований опубликованные данные представлены в таблице 1. В целом рост температур и концентрации СО2 способствует увеличению продуктивности.
Таблица 4.1 – Изменение продуктивности древесных растений, фитоценозов
Условия |
Регион |
Реакция |
Вид |
Источник |
1 |
|
2 |
3 |
4 |
Моделируемое глобальное изменение климата и удвоение концентрации CO2 |
(провинция Хэйлунцзян, Китай). |
Продуктивность древостоев, общий объем надземной биомассы и индекс листовой поверхности должны увеличиться соответственно на 7-28 %, 15-24 % и 5-8 %. |
Сосна корейская (Pinus karaiensis); ясень манчжурский (Fraxinus mandshurica); липа амурская (Tilia amurensis); орех манчжурский (Juglans mandshurica); бархат амурский (Phellodendron amurense); лиственница ольгинская (Larix olgensis); береза плосколистная (Betula platyphylla); тополь Давида (Populus davidiana). |
Chen, Wang, 2000 |
+2°C ;
удвоение концентрации CO2 |
Европа |
Может непосредственно влиять на деревья через: 1) снижение обмена через устьичный аппарат 2) усиление фотосинтеза.
В большинстве случаев это приведет к усилению прироста деревьев и эффективности водопоглощения. |
Guehl et al., 1999 |
|
|
|
|
|
|
|
|
|
|
|
|
|
Продолжение таблицы 4.1 |
1 |
|
2 |
3 |
4 |
+2СО2
+2°C |
Великобритания |
Большое, но кратковременное исходное повышение (на 20%) первичной нетто-продуктивности (NPP). |
|
Medlyn et al., 2000
|
|
Стойкое долговременное повышение NPP на 10-15%, рост минерализации N и поглощения N растениями. |
|
Текущие изменения за последние 50 лет |
Европа |
Отчетливо позитивные тенденции роста насаждений различного качества и лесорастительных условий в Центральной Европе и южных бореальных областях. В других регионах показатели не так отчетливы, однако снижение темпов роста наблюдается редко, что обусловлено местными факторами повреждений. Главными причинами ускорения роста лесов считают антропогенный привнос N, повышение концентрации CO2 в атмосфере, изменение климата |
Schuck, Karjalainen, Hunter, 2000 |
Текущие изменения за последние 50 лет |
Германия |
В последние 20 лет прирост насаждений разных пород увеличился на 12-43% по сравнению с таблицами хода роста. Вместе с тем отмечаются многочисленные случаи устойчивого снижения прироста при повреждении насаждений и стрессах (разные степени дефолиации). |
Von, 1999 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Продолжение таблицы 4.1 |
1 |
|
2 |
3 |
4 |
Повышение среднегодовой температуры на 1.1°С, среднегодовой минимальной температуры на 1.5°С, среднелетней температуры на 2.2°С и минимальной летней температуры на 2.3°С с 1950 по 1997 гг. |
Западная Франция |
С 1921 по 1991 гг. радиальный прирост увеличился на 45, 31 и 50% для ранней, поздней древесины и всего кольца соответственно.
Из-за негативной корреляции между летней температурой и шириной кольца повышение температуры не может объяснить наблюдаемое усиление роста.
Влияние азота (в среднем 6,3 и 11 кг N/га в год для общих и сквозных промывочных осадков соответственно); улучшение лесокультурной практики (посадка с более широкими междурядьями, более интенсивные прореживания) и удобряющее влияние CO2 рассматриваются как возможные причины наблюдаемых тенденций роста. |
Сосна корсиканская |
Lebourgeois et al., 2000 |
1981-1990 гг. |
Северное полушарие (45-70° с.ш.) |
Данные NDVI, полученные по снимкам со спутников NOAA, регистрируют увеличение продуктивности растительности за период 1981-1990 гг. |
Ваганов, Кирдянов, Силкин, 1999 |
|
Продолжение таблицы 4.1 |
1 |
|
2 |
3 |
4 |
+2СО2 |
США
Культивировали в пластмассовых контейнерах объемом 45 л с песчаной почвой с pH 5,1 и с очень низким содержанием доступных форм элементов минерального питания. |
При повышении концентрации CO2 наблюдалось усиление фотосинтетической активности, отмечалась тенденция уменьшения проводимости устьиц и повышения эффективности использования влаги. При высоких дозах N400 в сочетании с повышенной концентрацией CO2 усиливался рост и возрастал расход воды с одновременным повышением эффективности использования влаги. При низких дозах N20 удобрений отзывчивость на повышение концентрации CO2 слабая, расход воды меньше, ослаблялась острота проявления водного стресса. Максимальная интенсивность фотосинтеза при повышенной концентрации CO2 была в начале засушливого периода при высоких дозах азотных удобрений и несколько позднее - при низких дозах азотных удобрений. |
Сосна болотная (Pinus palustris Mill.) |
Runion et al., 1999
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Продолжение таблицы 4.1 |
1 |
|
2 |
3 |
4 |
Повышения концентрации атмосферного CO2 (700 ч. на млн) при неограниченной водообеспеченности и полном минеральном питании |
Австралия |
При повышенном уровне CO2 все виды за исключением медленнорастущей Acacia aneura увеличивали биомассу в среднем на 10% за 12 недель. Повышенная концентрация CO2 уменьшала площадь листьев на единицу их сухой массы через утолщение листовых пластинок. Интенсивность нетто-ассимиляции возрастала в среднем на 30%
Концентрация азота в растениях не была связана с ростом на повышенном фоне CO2 у медленно растущих видов акации, но снижалась на 10% у быстрорастущих видов. Скорость поглощения азота на единицу массы корней была выше у 7 видов при росте при повышенном уровне CO2, а площадь листьев на единицу массы корней уменьшалась у 7 из 10 видов. Абсолютный прирост RGR при повышенной концентрации CO2 был выше у быстрорастущих, чем у медленнорастущих видов акации. |
Acacia |
Atkin et al., 1999 |
|
|
|
|
|
|
|
|
|
|
|
Продолжение таблицы 4.1 |
1 |
|
2 |
3 |
4 |
Текущие изменения за последние 50 лет |
Калифорния, Невада, Аризона, Колорадо |
Значительное увеличение радиального прироста в течение последних 50 лет |
Pinus longaeva
Pinus flexilis |
Graybill, Idso, 1993 |
Текущая концентрация СО2 +300 ppm
в течение 23 месяцев |
Северная Америка |
Трехкратное возрастание скорости роста. |
Pinus eldarica |
Idso, Kimball, 1994 |
СО2 400 ppm – контроль; 800 ppm – опыт |
Северная Америка |
Тонкие корни (через 4 года) + 175%.
Ветви и ствол - через 5 лет +171 %; через 10 лет +107%.
Урожайность - +127%. |
Апельсиновые деревья |
Idso, Kimball, 1997 |
Потепление
+СО2 |
Северные районы Сибири и Дальнего Востока. |
Положительное влияние на рост лесов и накопление в них углерода. |
(Второе национальное сообщение..., 1998). |
+СО2 ;
увеличение суммы температур и средней температуры июля |
Западная Канада, северная Финляндия и Швеция |
Влияние температуры сильнее ощущалось в Швеции и Финляндии, чем в Канаде. Связи между 2 континентами различались на широте 56° с. ш. и южнее этой параллели. Плодородие почвы влияло меньше, но достоверно. |
Pinus contorta |
Fries et al., 2000 |
|
|
|
|
|
|
Продолжение таблицы 4.1 |
1 |
|
2 |
3 |
4 |
Изменения за период 1986-1999 |
Россия |
- урожайность практически всех культур возросла (Северный, частично Волго-Вятский, Уральский, Западно-Сибирский и Восточно-Сибирский районы);
- урожайность практически всех культур снизилась (Северо-Кавказский, Центрально-Черноземный, южные области Поволжского района);
Можно выделить три культуры, урожайность которых абсолютно возросла в России и в подавляющем количестве регионов - лен, картофель, кормовые травы.
Территориальный вектор увеличения урожайности направлен с юга на север в Европейской части и с запада на восток в Азиатской части России.
Рост урожайности культур с более низкотемпературным оптимумом роста и большим потреблением влаги (лен, картофель, кормовые травы). |
Сельскохозяйственные культуры |
Моисеев, Сухотин, 2000
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Продолжение таблицы 4.1 |
1 |
|
2 |
3 |
4 |
Моделируемое глобальное изменение климата и удвоение концентрации CO2 |
Китай |
Имитировали густоту смешанных древостоев, которая, согласно модельным оценкам, через 50 лет должна уменьшиться более чем на 20% в атмосфере. В древостоях вместо сосны корейской должны доминировать Quercus mongolica, Populus davidiana и Betula platyphylla. Дуб монгольский в общей биомассе древостоев должен занимать ~57%. При отсутствии климатических изменений продуктивность древостоя по прогнозам должна возрасти на 4% даже при условии преобладания в них Q. mongolica, P. davidiana и B. platyphylla. По данным модели общий индекс листовой поверхности древостоя также резко снижается |
Chen, Wang, 2000 |
Естественное повышение концентрации CO2 с 350 до 2600 мкмоль/моль |
Около термальных источников Италии, выделяющих CO2, |
Обогащение атмосферного воздуха CO2 до 750 мкмоль/моль увеличивает количество устьиц на единице поверхности листа, а от 750 до 2600 мкмоль/моль - увеличивает количество кутикулярного воска на листе. Смачиваемость листьев водой не зависит от концентрации CO2 в воздухе. |
Дуб каменный (Quercus ilex) |
Paoletti et al., 1998 |
На рисунке 22 приведена графическая интерпретация 279 экспериментальных исследований, в которых растения всех типов выращивались в условиях стресса (пустые кружки) и без стресса (жирные кружки). больший стимулирующий эффект рост СО2 производит на растения, произрастающие в более засушливых (стрессовых) условиях.
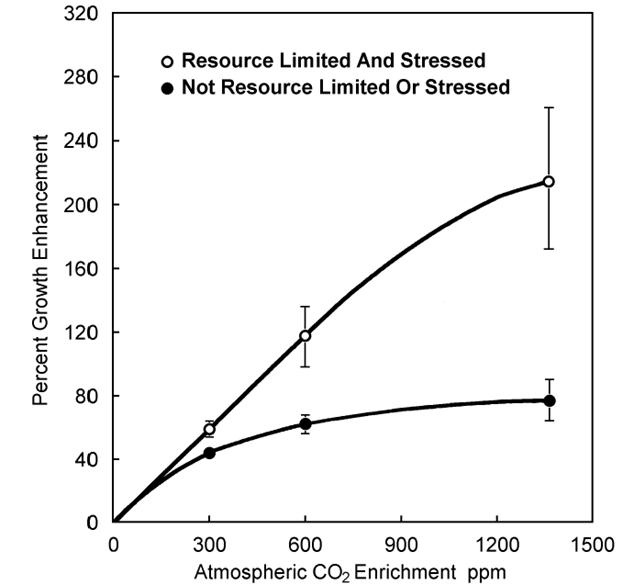
Рисунок 4.1 - Обобщенные данные 279 экспериментальных исследований влияния увеличения содержания СО2 в атмосфере(Robinson et al., 1998).
Изменения климата воздействуют на частоту, интенсивность, время и/или пространственную локализацию потрясений. Многие потенциальные последствия будущих климатических изменений могут быть смягчены приспособлением лесных сообществ к естественным вариациям климата. Тем не менее, масса исследований свидетельствует о том, что, в результате возникновения новых режимов воздействия при изменении климата возможны существенные изменения в лесах с долговременными экологическими и социально-экономическими последствиями.
Природные потрясения накладываются на последствия человеческой активности, такие как загрязнение воздуха, выращивание и сбор урожая, сельскохозяйственное и городское строительство, отдых на природе. И речь не идет об обычном суммировании.
Естественные воздействия, связанные с климатом, проистекают от насекомых, болезней, привнесенных видов, пожаров, засух, обильных снегопадов, ураганов, оползней, штормового ветра и обледенений. В каждую геологическую эпоху местные, региональные и глобальные изменения температуры и режима осадков влияют на частоту и интенсивность такого рода природных факторов.
Влияние этих потрясений рассматривается в широком спектре шкал, от листа и дерева до леса и лесного ландшафта. Их результат может проявляться в: утрате листвой цвета и сокращения периода его функционирования; деформации структуры дерева – ветровал, ветролом, сломанные ветви или потеря кроны; росте смертности деревьев или хронический стресс, приводящий к их гибели; дисфункции регенерации – утрате семенного фонда; деградации физического окружения, включающего эрозию почв; дисбаланс в обороте биомассы и питательных элементов; воздействии на поверхностные органические слои почвы и подземные растительные корневые и репродуктивные ткани; и нарастающая гетерогенность ландшафта (разделенность, нарастание свободных пространств между лесными сообществами).
Внедренные (инвазивные) виды могут влиять на лесные экосистемы посредством питания (травоядность или хищничество), разрушения среды обитания, конкуренции, утраты генетического фонда при скрещивании с местными видами и болезней (привносимых или приобретаемых). Вспышки местных болезней или численности насекомых могут повлечь сходные последствия для леса. Хотя леса в США основательно охраняются, всего пять видов насекомых поражают около 21 миллиона акров каждый год (Designing a report..., 1999).
На уровне экосистемы привнесенные виды – так же, как и вспышки численности местных насекомых, эпидемий – могут менять природные циклы и режим природных воздействий – такие как циклы питания, частота и интенсивность пожаров (Mack, D’Antonio, 1998.). Некоторые виды деревьев приспособились выживать в периоды повторяющихся потрясений. Толстая кора отдельных пород (сосна обыкновенная, лиственница) позволяет им переживать низовые пожары. Повторяющиеся низовые пожары сокращают плотность подроста, который может служить горючим материалом для перехода огня на кроны. Таким образом, повторяющиеся низовые пожары сокращают частоту убийственных верховых пожаров.
Для экосистем бореальных лесов предполагается, что влияние климатических изменений будет опосредовано в изменениях частоты пожаров, динамики прогалин, на доминанты вредителей (Bergeron et al., 1998).
Одни влияния накладываются на другие, появление одного фактора стимулирует появление и развитие другого, часто более опасного. Например, засуха часто ведет к всплеску численности насекомых, болезням или пожарам. Насекомые и болезни также создают большие «запасы горючих материалов» и поэтому способствуют учащению пожаров.
Отмечаемый в северном полушарии (и соответственно в Сибири) доминирующий рост температуры воздуха в зимнее время, сопровождаемый оттепелями и снегопадами, будет способствовать снижению устойчивости древесных растений, а также более благоприятной зимовки насекомых. Следствием сочетания снегопадов с оттепелями будут интенсивные снеголомы (особенно в лесных культурах). Насекомые и пожары завершат процесс разрушения.
Многие лесные регионы подвергались воздействию человека в прошлом и сегодня ситуация здесь отлична от прежней. Например, борьба с пожарами в приспособленных к пожарам западных лесах США привела к нарастанию густоты леса и биомассы, изменению состава леса, а также способствовала нарастанию болезней, вспышек численности насекомых (Flannigan et al., 2000; Ayres, Lombardero, 2000).
Некоторые типы лесов адаптировались к периодическим потрясениям и даже стали зависеть от них. Леса длиннохвойной сосны (Pinys Longifolia Roxb.), например, часть такой «пожарозависимой» экосистемы, которая не могла бы существовать, если бы пожары не были повседневны для Юго-востока. Около 1,5 миллионов акров леса было целенаправленно сожжено (а также были применены и другие формы сокращения массы горючего материала) в 1998 для того, чтобы улучшить здоровье и видовое разнообразие леса путем восстановления «пожарозависимых» экосистем, пострадавших от долговременной борьбы с пожарами (USDA Forest Service, 1998).
Следует учитывать, что длительное действие (в течение многих поколений) одного какого-либо фактора в запороговых значениях обеспечивает выживание лишь тем особям, в генотипе которых произошли изменения, направленные на специфическую адаптацию к лимитирующему фактору, что также сопровождается снижением продуктивности, но, в отличие от первого случая, необратимо при снятии действия лимитирующего фактора. Это необходимо учитывать при оценке возможных последствий глобального изменения климата: нужно быть готовыми к искусственному возобновлению древесной растительности в тех регионах, где произошли глубокие адаптивные изменения в метаболизме древесных растений, особенно на вечной мерзлоте (Милютина, Судачкова, Семенова, 1998).
Усиливающиеся в последние два столетия антропогенные нарушения естественных взаимодействий бореальных лесов и климата в значительной мере способствовали тому, что на огромной территории хвойные леса сменились листопадными. Это изменение ландшафта связывается с происшедшими на бореальных широтах изменениями температуры, фотосинтетической активности, содержания CO2 в атмосфере, высоты весеннего снежного покрова (Kronberg, Watt, 2000). С точки зрения авторов, такие признаки указывают на уменьшение способности бореальных ландшафтов сохранять вещество и энергию и участвовать в их кругообороте. Наряду с климатическими изменениями (безусловно оказывающими существенное влияние) исключительно велика роль хозяйственной деятельности человека (рубка леса, сельскохозяйственное производство и пр.) При изучении характера и изменения лесов на юге Швеции на протяжении последних 3000 лет (пыльцевой анализ), установлено что быстрее всего растительность менялась в последние 150 лет (кульминация изменений, начавшихся 850 лет назад) (Lindbladh, Bradshaw, Holmqvist, 2000). В качестве генеральной линии развития установлен переход от богатого смешанного листопадного леса (Alnus, Corylus, Quercus и Tilia через Betula и Carpinus) к хвойному (в основном Picea и Pinus) и редко к лесу из Fagus. Медленное уменьшение роли доминантов прежних лесов - Corylus, Quercus и Tilia началось ~700 г. н. э. и сопровождалось увеличением отложения пыльцы хлебных злаков, что свидетельствует о деятельности человека, как об основной движущей силе этих процессов.
Предполагаемое в будущем дальнейшее потепление наращивает опасность пожаров, даже в самое ближайшее время. Одновременно, вероятность возникновения лесных пожаров возрастает в связи с резким увеличением числа посещений лесов людьми.
Биомасса сибирских лесов сосны обыкновенной (лишайниковые и мохово-брусничные) сильно зависит от пожаров, а изменение климата включается в экосистемные функции преимущественно через изменения пирологического режима (Wirth et al., 1999)
Частота лесных пожаров в северной Азии, как ожидается со средней степенью вероятности, возрастет (Technical Summary Working Group II..., 2001). Температура воздуха у поверхности воды, в частности, может послужить фактором, благоприятствующим возникновению молний, которые чаще будут поджигать лес. Лесные пожары, как ожидается, будут случаться чаще в северных частях бореальной Азии в результате глобального потепления.
Установлено быстрое изменение режимов пожара в ответ на изменение климата (Flannigan et al., 1998; Stocks et al., 1998), так что эта реакция имеет шанс превзойти по своему влиянию прочие факторы, связанные с влиянием изменения климата на распределение видов, их миграцию или исчезновения в пораженных огнем зонах.
В анализе Фланнигана и др. (Flannigan et al., 2000) предполагается, что разрушительная сила будущих пожаров будет возрастать на большей части территории Северной Америки под действием как изменений климата, так и ряда других факторов (данные для США и Канады существенно варьируют). Более теплый и влажный сценарий «Хадли» предполагает возрастание числа и силы пожаров примерно на 20% для Северо-Востока с некоторым сокращением для северной части Великой Равнины, с нарастанием менее чем на 10% для остальной части континента. Теплый и сухой «Канадский» сценарий предсказывает 30%-е возрастание разрушительной силы пожаров для Среднего Запада, Аляски и отдельных регионов Юго-Востока, и примерно 10%-й рост в прочих частях континента. Этот же прогноз предсказывает увеличение площади выгорания на 25-50% в пределах США.
Температура и осадки – не единственные факторы, влияющие на режим пожаров; например, по оценкам общей циркуляционной модели Института пространственных исследований Годдарда и основанного на ней сценария частота молний возрастет на 44% (Price and Rind, 1994). Другие факторы, такие как длительность пожароопасного сезона, природные условия после загорания, и характеристики растительности также оказывают воздействие на режим пожара.
Утверждается, что пожароопасный сезон продлится в среднем на 30 дней дольше в условиях удвоенного содержания в воздухе углекислоты, - по сравнению с современными климатическим условиями Канады (Wotton and Flannigan, 1993). Ущерб, наносимые лесными пожарами был также чувствителен к изменениям ветра, как и к изменениям температуры и в выпадении осадков - в анализе чувствительности к изменениям климата, осуществленном для Калифорнии (Torn and Fried, 1992). Человеческая деятельность – борьба с огнем и землепользование – также будут влиять на режим пожаров в будущем. Например, вредоносность лесных пожаров на территориях западной части США выросла в 1980-х годах после 30 лет интенсивной борьбы с огнем, приведшей к возрастанию лесной биомассы и отпада (Keane et al., 1998).
Другое модельное исследование было сфокусировано на влиянии изменений климата на растительность во взаимодействии с лесными пожарами (Bachelet et al., 2000). Количество сгоревшей биомассы при изменениях климата в будущем в динамической глобально-вегетативной модели (МС1) возрастает. В этой модели, возникновение, разрушительная сила и размер пожара представлены как прямая функция природных условий и горючего материала (Lenihan et al., 1997; Daly et al., 2000; Bachelet et al., 2000). В западной части США возрастающая температура, стабильные или нарастающие осадки, распространение избыточного СО2 и рост эффективности использования воды улучшают продуктивность экосистемы, что приводит к увеличению биомассы. Значительные вариации климата (сухие годы сменяются влажными) и прирост горючего материала вызывают рост количества и интенсивности пожаров.
Во всех прогнозах предсказывают расширение площади, поражаемой огнем с изменением климата. Эти исследования основаны на физических и биологических факторах, определяющих потенциальную угрозу пожара. Такие факторы, как текущее управление земельными угодьями, землепользование, и формы собственности во внимание не принимались. Сельскохозяйственная деятельность и использование лесных территорий в иных целях могут также оказать влияние на количество и вид горючего материала.
Подтверждением сказанному является современная (на 21 июня 2002 г.) катастрофическая ситуация в США. Площадь пожара Хеймэн достигла 55 тысяч га. Эвакуировано около 9 тысяч человек. Площадь пожара около Дюранго достигла 24 тысяч га. Всего в штате Колорадо пройдено огнем 80 тысяч га. Разрушено 140 домов. Гораздо более катастрофическая ситуация складывается в Аризоне. Площадь пожара на востоке этого штата достигла 51 тыс. га. Пожарные опасаются, что он может слиться в другим, быстро увеличивающимся соседним очагом огня, площадь которого пока только 6 тысяч га. Огнем пройдено 800 тысяч га, что в два раза выше средних значений за последние 10 лет.
Засухи случаются практически во всех лесных экосистемах (Hanson and Weltzin, 2000). Первая немедленная реакция деревьев на засуху заключается в сокращении общей первичной продукции (NPP) и использовании воды, обуславливаемая уменьшением влажности почвы и устьичной проводимости. При продолжительной суровой засухе растения гибнут. Саженцы и побеги обычно гибнут первыми, уступая даже умеренной засухе. Сильная корневая система, запас карбогидратов и питательных элементов в крупных деревьях делают их чувствительными только к продолжительной, суровой засухе. Имеют место и вторичные эффекты. Там, где сокращение в NPP значительно или продолжается в течение нескольких последовательных сезонов, возможно нарастание чувствительности к насекомым и болезням, особенно в густых лесных массивах (Negron, 1998). Засуха также может тормозить процессы разложения, что ведет к накоплению органических веществ в лесной подстилке.
Последствия изменения режима засух зависят от годовых и сезонных изменений климата и от того, усиливает ли текущая адаптация растения к засухе гибкость и сопротивляемость новым условиям. В лесах есть тенденция к достижению максимальной площади листа, при которой используется практически вся имеющаяся в наличии в почве влага (Eagleson, 1978; Hatton et al., 1997; Kergoat, 1998; Neilson and Drapek, 1998). Даже небольшое повышение температуры в сезон роста может ускорить испарение и привести к водному стрессу. Основываясь на этом предположении, результатах MAPPS, биогеографическая модель и MCI (динамическая глобально-вегетативная модель) – подтверждают вывод о том, что нарастание испарения могут в будущем усилить воздействие засухи на леса США (Юго-Восток, южная часть Скалистых гор и часть Северо-запада) (Neilson and Drapek, 1998; Bachelet et al., 2000).
Современное поколение экологических моделей (MAPPS, MC1 и др.), учитывают механизмы адаптации леса к засухе, такие как изменение скорости роста или подземно-надземное накопление углерода. Эти приспособления смягчают воздействие климатических факторов – в том числе и засухи – на индивидуальные деревья и лесные массивы в целом.
Если климатические изменения интенсифицируют все конвективные процессы в атмосфере, то они будут и увеличивать частоту и интенсивность ветров, в т.ч. торнадо и ливневых дождей (Berz, 1993). Карл и др. (Karl et al. (1995a) обнаружили, что доля осадков, выпавших в экстремальных природных ситуациях, возросла в США с 1910 по 1990 годы. Карл и др. (Karl et al. (1995b) далее предполагают, что климат США стал более экстремальным (в терминах аномалий температуры и осадков) в течение последних десятилетий.
Климатические потрясения часто приводят к ущербу на значительных территориях. Например, 25 октября 1997 года шторм повредил более 13 тыс. акров елового леса в штате Колорадо (USDA Forest Service Routt National Forest, 1998), а шторм 4 июля 1999 года – около 250 тыс. акров леса в штате Миннесота (Minnesota Dept. of Natural Resources press release, 7/12/99).
Климат, так же как и человеческая деятельность, в значительной степени определяет потенциал и уровень распределения интродуцированных видов (Simberloff, 2000). Неподходящий климат в точках внедрения ухудшает условия выживания значительного большинства интродуцированных видов (Williamson, 1999). В более теплых частях США интродуцированные виды составляют существенную часть биоты (Simberloff, 1997). Там, где холодный и сухой климат сдерживает проникновение, изменения в температуре и осадках могут привести к ускоренному росту, размножению и расширению их ареалов. Например, лабораторные исследования Adelges piceae в различных температурных условиях, дали основу для предположений о том, что происходящие под влиянием температуры изменения популяционной динамики насекомого воздействуют на выживание Abies fraseri (Dale et al., 1991).
Ключевая черта большинства интродуцированных видов состоит в том, что они способны выжить в неблагоприятных условиях по причине высоких репродуктивных возможностей, хорошей способности к рассеянию и быстрого роста (Vitousek et al., 1996). Если изменения климата влекут за собой нарастание потрясений – таких, как пожары или засуха, эти экосистемные стрессы создают именно такой тип окружения, в котором интродуцированные виды могут быстро распространяться. Взаимодействие между интродуцированными видами, автохтонными сообществами, человеческой деятельностью, фрагментирующей экосистемы, нарастающими перемещениями атмосферного воздуха и изменениями климата может положительно воздействовать в смысле доминации пришельцев, но предсказание специфических последствий интродукции остается проблематичным (Dukes and Mooney, 1999; Williamson, 1999).
Вспышки численности насекомых и патогенных микроорганизмов могут существенно влиять на природную среду, природные биоценозы, производство древесины и экологические процессы. Это особенно важно в связи с большей восприимчивостью насекомых к внешним воздействиям в силу их меньшей инерционности по сравнению с фитоценозом (Исаев и др., 1999).
Нынешнее потепление уже сопровождается потрясениями лесов, проистекающими от насекомых, штормовых ветров и пожаров. Распространение большого елового короеда (Ips typographus L.), в прошлом ограниченное по причине холодов, с 1992 года привело к массовой гибели деревьев на более чем 0,93 млн. га Кенайского полуострова (Designing a report..., 1999). В то же самое время, нарастание гибели деревьев при интенсивных штормах и повреждений лесного полога снежной массой, типичное для теплых зим, может повысит чувствительность лесов к нападению насекомых.
В период с 1986 по 1995 годы в США четыре местных вида насекомых ежегодно поражали следующие площади: более 1,62 млн. га - западная еловая листовертка; менее одного миллиона акров – восточная еловая листовертка; менее двух миллионов акров – лубоед желтой сосны; и около 4,86 миллионов га – лубоед сосновый (Designing a report..., 1999). В то же самое время, площадь, пораженная одним из этих видов, изменялась от мене чем половины от долговременной средней величины до трехкратного ее превышения. Около 5,26 миллионов га южных лесов было подвержено воздействию одного заболевания - Cronartium fusiforme, и 11,74 млн. га западных лесов поражено растением-паразитом, карликовой омелой (Designing a report..., 1999). Потрясения, такие как засуха или пожары, влияют на эти вспышки.
Масса научной литературы по предмету представляет нашему вниманию множество процессов, где возрастающие запасы углекислоты в атмосфере и изменения климата могут существенно менять характер воздействия на лес со стороны насекомых и патогенов (Ayres and Lombardero, 2000). Накопление углекислого газа и изменения климата могут как усилить, так и ослабить воздействие патогенов и насекомых посредством прямого влияния на выживание, размножение и расселение этих организмов. Например, нарастание межгодовых вариаций минимальных зимних температур, возможно, благоприятствует самым северным вспышкам численности соснового лубоеда, препятствуя, в то же время, аналогичным вспышкам на юге (Ungerer et al., 1999). Также представляется возможным, что изменения климата будут влиять на механизм насекомо-патогенных воздействий непрямым образом, изменяя численность их естественных врагов и соперников. Кроме того, климат и углекислота влияют на восприимчивость деревьев к насекомым и патогенам посредством изменений в химии растительных тканей (Ayres and Lombardero, 2000). В листьях растений, сформировавшихся в условиях высоких концентраций углекислого газа, меньше содержание азота, а следовательно, белка (Алексеев и др., 1999). Поэтому ценность такой растительности как пищи для животных и насекомых значительно снижена. Действительно, в экспериментах насекомые начинали съедать существенно больше листьев этих растений по сравнению с растительностью в условиях нормальной концентрации СО2. Причем у насекомых замедляется развитие, они становятся более уязвимыми для хищников и паразитов.
Короткие жизненные циклы, высокая подвижность, репродуктивный потенциал, и физиологическая чувствительность к температуре приводят к тому, что даже незначительное изменение климата может оказать существенное влияние на распространение и численность многих лесных насекомых и патогенов. Благоприятное воздействие может иметь место там, где тонкий слой снега приводит к усиленной зимней смертности растений, вредное же – когда потепление ускоряет развитие насекомых и способствует распределение насекомых и патогенов в районах, где низок уровень сопротивления деревьев. Вредные эффекты также могут быть следствием наложения многих факторов. Например, потепление может усиливать вспышки численности насекомых в бореальных лесах, что, в свою очередь, ведет к учащению пожаров (Ayres and Lombardero, 2000). Уже сейчас достаточно широко распространены эффекты, связанные с болезнями и насекомыми. В 1995 году более 36 млн. га лесной площади в США были поражены всего несколькими болезнями и вредителями: еловой листоверткой (западной и восточной), лубоедом сосновым и лубоедом желтой сосны, корневой губкой и ржавчинным грибом Cronartium fusiforme (Ayres and Reams, 1997; Ayres and Lombardero, 2000).
Потенциальная опасность для сосновых лесов от сосновой пяденицы в ряде районов Центральной Сибири возникает при повышении температуры на 2-4°С (Исаев и др., 1999). Анализ показал, что при рассмотрении влияния глобальных изменений климата на динамику численности популяции насекомых в рамках такого подхода недостаточно иметь данные о средних значениях климатических сдвигов. Необходимо знание особенностей климатических изменений в течение сезона, поскольку в анализированной модели различные режимы динамики численности реализуются при одних и тех же средних значениях климатических параметров, но различных их дисперсиях. Так, например, для имитационной модели сосновой пяденицы зависимость динамики численности от дисперсии климатических показателей немонотонна. Для сценариев со значительным ростом средних температур и осадков увеличение дисперсии этих показателей сопряжено с уменьшением вероятности вспышек вредителя. Для сценариев с небольшим увеличением климатических показателей вероятность возникновения вспышек с ростом дисперсии возрастает. В сценариях со средними значениями климатических показателей динамика численности вредителя практически не зависит от дисперсии (Исаев и др., 1999).
|


