|
УДК 630*5: 630*443.3
Павлов И.Н., Барабанова О.А., Миронов А.Г.
Распределение деревьев сосны обыкновенной по диаметру в очагах интенсивного биогенного воздействия
Представлены результаты исследования распределения деревьев сосны обыкновенной по диаметру в очагах интенсивного биогенного воздействия. Наряду с корневыми патогенами (Armillaria mellea sensu lato) в исследуемых насаждениях были отмечены высокие уровни зараженности деревьев раком-серянкой (Cronartium flaccidum (Alb. et Schw.) Wint.; Peridermium pini (Willd.) Kleb.) и заселенности Blastophagus minor Hart. В процессе усыхания образуются очаги диаметром 10-30 м, в дальнейшем сливающиеся в полосы. Несмотря на одновозрастность древостоев сосны, в распределении, как живых, так и усохших деревьев имеет место две вершины. Успешная аппроксимация оказалась возможной только с использованием многокомпонентной смеси нормальных распределений. Первая составляющая (Dср живых = 15,4 см) образована в результате отсутствия своевременного отпада отстающих в росте деревьев (по генотипу или произрастающие на маломощных почвах). Среди них и образуется первая составляющая сухостоя со средним значением диаметра 11 см. Отпад происходит естественно (действие патогенов вторично) в результате угнетения со стороны соседних деревьев. В дальнейшем именно на их основе происходит усиление вирулентности и агрессивности опенка. В итоге, вторая составляющая отпада (Dср = 21 см) образована действием опенка, как первичного патогенна, что способствует рассечению общего распределения живых деревьев на две составляющие.
Распределение числа деревьев по диаметру является важным показателем, характеризующим состояние древостоя. На него оказывает влияние большое количество факторов: возраст, условия произрастания, породный состав, полнота, характер размещения, антропогенное и биогенное воздействие, пожары, ветровалы и пр. Форма кривой распределения диаметров определяется видом и интенсивностью естественного изреживания и является его характеристикой (Свалов, Свалов, 1973). Определенную закономерность в возрастной динамике рядов распределения диаметров стволов отмечают А. А. Макаренко (1975); Н. Н. Свалов, С. Н. Свалов (1973); В. В. Кузьмичев, Ю.Н. Савич (1979); В.Я Каплунов (1989); А. И. Бузыкин и др. (2002).
Согласно современным представлениям о гомеокинезе биологические системы стремятся не к абсолютной стабилизации своего состояния, а к удержанию его в определенных пределах при непрерывном колебательном изменении параметров вокруг определенных точек равновесия (Свирежев, Логофет, 1978). Одним из механизмов поддержания стабильности в экосистеме является организация отрицательной обратной связи (Одум, 1986). На разных возрастных этапах максимальный запас древостоя формируется различным количеством деревьев. Естественный отпад не только удаляет из древостоя генетически худшие экземпляры, но и создает структуру, обеспечивающую устойчивость ценоза. В свою очередь, загущенность среди деревьев нижних ступеней толщины и, как следствие, высокая положительная асимметрия выполняют роль обратной связи, определяющей, какое количество деревьев необходимо элиминировать из древостоя.
При определенном сочетании факторов (высокое плодородие почвы в сочетании с малой мощностью; равномерное размещение возобновления и его одновозрастность) происходит нарушение своевременного отпада. При произрастании леса на маломощных дерновых почвах, подстилаемых на глубине 20-30 см твердыми горными породами, корневая система сосны развивается в верхнем, легкопересыхающем слое (исследуемое нами насаждение). В итоге стержневой корень отсутствует или сильно редуцирован. До определенного времени насаждение развивается как высокопродуктивное. В дальнейшем после превышения эдафического потенциала поддерживать необходимую потребность насаждения в элементах питания и влаге наступает снижение устойчивости фитоценоза в целом. В первую очередь страдают деревья, располагающиеся в условиях жесткой конкуренции, на маломощных почвах и с плохо развитой корневой системой. Особую опасность для них представляет наступление даже кратковременного засушливого периода. При этом отрицательные последствия могут не проявляться длительное время. Как правило, недостаток влаги в почве ведет к потере части ассимиляционного аппарата, тонких корней и как следствие - снижение устойчивости к болезням и вредителям. При этом с момента заражения до гибели может пройти достаточно длительный промежуток времени. Анализ годичных колец 80-110-летних погибших хвойных показал, что деревья были инфицированы дереворазрушающими грибами комплекса Armillaria mellea sensu lato 50 лет назад (Shaw, Kile, 1991).
Внедрение столь сильного патогена безусловно не может не сказаться на строении древостоя. Чаще всего подобное воздействие ведет к усложнению структуры, требующей для своего описания оригинальных функций.
Для описания случайных величин в прикладных исследованиях используется около 30 законов и семейств распределений (Лемешко, 1998). Естественно, что это не покрывает существующего многообразия распределения случайных величин. Простые модели часто не могут описать рост числа случайных величин, возникающих на «хвостах» распределений, которые определяются другими параметрами (например, почвенная мозаичность; развитие болезней и вредителей, экстремальное климатическое воздействие).
Сложные биологические, технические, социальные системы включают совокупности элементов, принадлежащих, как правило, нескольким классам. Примером такой совокурности является система, состоящая из нормально функционирующих (1-й класс) и ослабленных (2-й класс) объектов (элементов). Каждый класс объектов можно охарактеризовать некоторой функцией плотности распределения, описывающей закон распределения анализируемого признака. Распределение в такой системе может быть унимодальным, бимодальным и полимодальным.
Программа и методика исследований
В нижнем течении р. Караульной (ООПТ зеленая зона г. Красноярска) на прилегающих сопках в последнее десятилетие наблюдается интенсивная куртинная гибель сосны обыкновенной. Усыханию подвержены деревья всех классов Крафта (рис. 1). Погибают экземпляры, не испытывающие каких-либо признаков угнетения, с хорошо развитой кроной. Гибель наступает очень быстро. Образуются очаги диаметром 10-30 м, в дальнейшем сливающиеся в полосы. Часто наблюдается гибель деревьев по периметру существующих окон в древостое. Куртинное усыхание, наличие плодовых тел Armillaria mellea sensu lato, а также ряд других диагностических признаков (Павлов, Миронов, 2004) свидетельствуют об интенсивном развитии армилляриоза. Исключительно быстрая гибель отдельных деревьев сосны, ослабленных корневым патогеном, вызвана их интенсивным заселением малым сосновым лубоедом. Для усохших деревьев также характерна более высокая частота встречаемости рака-серянки.
Для изучения кольцевых процессов распространения агрессивных видов грибных патогенов на полигоне 10,8 га было заложено 13 круговых ППП общей площадью 1,17 га. Использование круговых постоянных пробных площадей с картированием из центра через измеряемые угол и расстояние (Павлов, 2005) имеет значительные преимущества в сравнении с методами, предполагающими разбивку исследуемой территории регулярной сеткой. При значительном снижении трудоемкости сохраняется высокая точность определения координат. Исследуемый древостой сосны обыкновенной (9С1Ос + Б ед. Лц,) 5 класса возраста имеет полноту 1,0 (2 бонитет).
Изучение изменения в строении древостоев в условиях интенсивного биогенного воздействия представляет не только теоретический, но и практический интерес.
Впервые алгоритм разделения совокупности на две нормальные составляющие методом моментов предложил К. Пирсон. В дальнейшем метод был упрощен, появились работы по разделению смеси биноминальных распределений, Пуассона, Вейбулла (Никитин, Швиденко, 1978).
Уравнение функции плотности смеси вероятностных распределений f(x), описывающее закон распределения анализируемого признака во всей (объединенной) генеральной совокупности имеет вид:
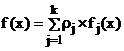 , (1) , (1)
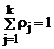 , rj ? 0, , rj ? 0, 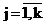 , (2) , (2)
где rj – удельный вес (доля) j - го компонента смеси (априорные вероятности появления наблюдений из j - го класса объектов), fj(x) = jj(aj,x) - плотность распределения анализируемого признака для j- го класса, aj – векторные параметры, от значений которых зависит закон распределения j- го компонента смеси, 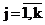 - номер класса, k – число классов. - номер класса, k – число классов.
Результаты и обсуждение
Апробировано шесть чаще всего применяемых распределений (нормальное, бета, Вейбулла, логнормальное, Пуассона, экспоненциальное). Насколько хорошо выборка описывается теоретическим законом, проверяли с использованием различных критериев согласия (Колмогорова, Лемана-Розенблата, Пирсона). Основная идея критериев согласия состоит в измерении расстояния между функциями эмпирического и теоретического распределения (Орлов, 2002).
В качестве контроля было выбрано сходное по таксационно- лесоводственным показателям насаждение без признаков заболевания, примыкающее к древостою с признаками армилляриоза (рис. 2).
Асимметрия распределения по диаметру живых деревьев составляет 0,17±0,09. Гипотеза о нормальности представленного распределения отклонена на основании критерия Шапиро-Уилкса (p=0,00008). По критерию Колмогорова-Смирнова гипотеза была отклонена при несколько меньшем уровне вероятности (p<0,1).
Асимметрия распределения по диаметру сухостоя составляет 0,45. Однако гипотеза о нормальности распределения не может быть отклонена на основании критериев Шапиро-Уилка (p=0,42) и Колмогорова-Смирнова (p> 0.20). Причина противоречия заключается в малом числе наблюдений.
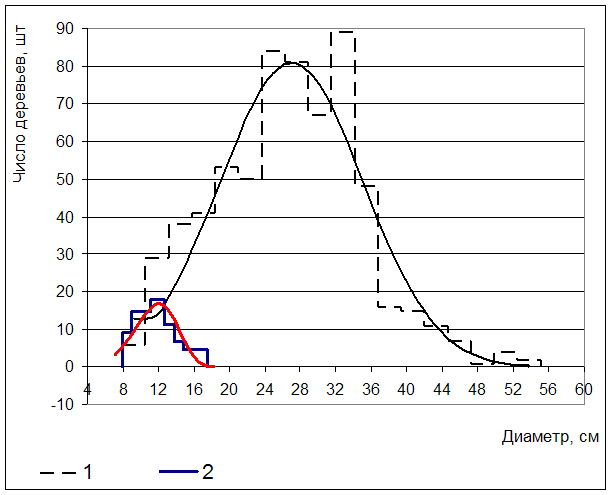
Рис 2. Распределение деревьев сосны обыкновенной в контроле (1- живые; 2 – усохшие)
Для нарушенных древостоев, при проверке согласия (по критериям Колмогорова, Лемана-Розенблата, Пирсона) по значениям достигнутого уровня значимости, ни одна из функций не может быть принята.
Поэтому для аппроксимации было решено воспользоваться многокомпонентной смесью нормальных распределений:
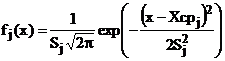 , , 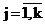 , (6) , (6)
где Xсрj , Sj2 – оценки математического ожидания и дисперсии j – й компоненты к компонентной функции плотности смеси нормальных распределений древостой сосны обыкновенной по диаметру х.
В табл. 2 представлены результаты проверки согласованности эмпирической и теоретических функций смесей распределений по критериям согласия Колмогорова (D), Лемана – Розенблатта (w2), Пирсона (c2). Учитывая рекомендации (Орлов, 2002; Справочник по …, 1989, 1990) проверку согласованности функций распределения проводили при уровнях значимости a1 = 0,2 – для критерия Колмогорова, a2 = 0,5 – для критерия Лемана – Розенблатта, a3 = 0,05 – для критерия Пирсона.
Таблица
Результаты согласованности эмпирических и расчетных функций распределения по критериям согласия
Проверка гипотезы о соответствии функции распределения выборочных данных с функциями смесей нормальных распределений |
Критерий
согласия |
Показатели |
Число компонентов нормального распределения |
живые деревья |
сухостой |
1 |
2 |
1 |
2 |
Колмогорова
(D) |
Dрас |
0,0378 |
0,0206 |
0,1468 |
0,0373 |
a |
0,200 |
0,200 |
0,200 |
0,200 |
Dkp(a) |
0,0440 |
0,0440 |
0,0879 |
0,0879 |
Вывод: гипотеза |
не отклоняется |
не отклоняется |
отклоняется |
не отклоняется |
Лемана-Розенблатта (w2) |
(Nw2)рас |
0,236 |
0,047 |
0,635 |
0,033 |
a |
0,500 |
0,500 |
0,500 |
0,500 |
(Nw2)кр |
0,118 |
0,118 |
0,118 |
0,118 |
Вывод: гипотеза |
отклоняется |
не отклоняется |
отклоняется |
не отклоняется |
Пирсона (c2) |
cpac2 |
42,555 |
20,458 |
68,634 |
15,458 |
число параметров, с |
2 |
5 |
2 |
5 |
Число степеней свободы |
15 |
12 |
15 |
12 |
a |
0,050 |
0,058 |
0,050 |
0,217 |
ckp2 |
24,996 |
20,512 |
24,996 |
15,465 |
Вывод: гипотеза |
отклоняется |
не отклоняется |
отклоняется |
не отклоняется |
На рисунке 3 представлено распределение по диаметру деревьев, произрастающих в очагах заболевания, с выделением живых и сухостойных.
Погибшие деревья образуют ряд распределения с двумя вершинами. Данная закономерность имеет биологическую интерпретацию. Экземпляры, образующие первое нормальное распределение со средним значением диаметра 11 см, усыхают естественно (действие патогенов вторично) в результате угнетения со стороны соседних деревьев. Доля первой составляющей (?) несколько меньше второй (0,42). В дальнейшем усиление вирулентности и агрессивности опенка связано с его функционированием на фоне ослабления биоценоза в результате высокой рекреационной нагрузки, техногенного воздействия, вырубки единичных деревьев и значительным естественным возрастным отпадом осины и березы. В итоге вторая составляющая отпада (Dср = 21 см) образована действием опенка, как первичного патогена.
Асимметрия распределения по диаметру составляет 1,09±0,19. Гипотеза о нормальности представленного распределения отклонена на основании критериев Шапиро-Уилкса (p=0,0000) и Колмогорова-Смирнова (p<0,01).
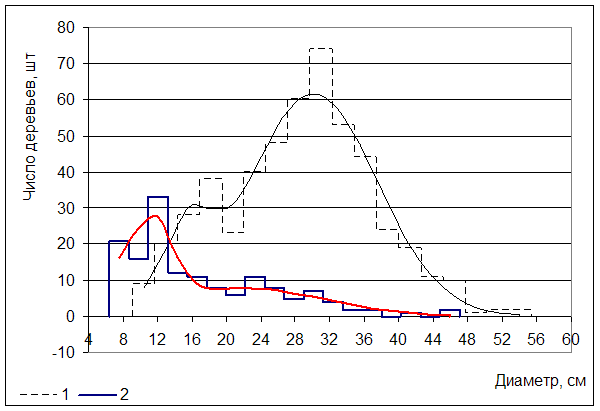
Рис. 3 Распределение числа деревьев по диаметру (1- живые; 2 – усохшие)
Живые деревья, произрастающие на круговых площадках в очагах усыхания, также образуют ряд распределения с двумя вершинами. Первая составляющая (Dср = 15,4 см) образована угнетенными деревьями (доля числа наблюдений – 14,5 %), вторая (Dср = 30 см) – состоит из особей входящих в основной полог. Деление на два класса вызвано патогенным действием опенка. Вторая вершина сухостоя (Dср=21 см) свидетельствует о направлении и объектах воздействия биотрофов, что способствует рассечению общего распределения живых деревьев. Асимметрия распределения живых деревьев по диаметру составляет 0,01. Несмотря на столь близкое значение к нормальному распределению, критерии Шапиро-Уилкса (p=0,0003) и Колмогорова-Смирнова (p<0,01) не подтвердили гипотезу о нормальности представленного распределения.
Заключение
Наличие двухвершинного (и более) распределения является закономерным в разновозрастных многоярусных насаждениях. В описываемых случаях речь идет об одновозрастном насаждении сосны обыкновенной с примесью березы и осины. Для одновозрастного древостоя светолюбивой сосны это не является типичной формой. И действие патогенов является следствием избыточной густоты среди деревьев из низших ступеней толщины и направлено на формирование более симметричного распределения.
Е.Л. Маслаков (1984) указывает, что ранговая структура популяций с возрастом насаждений меняется мало. Относительное сходство в строении древостоев было выявлено еще работами Фекете, Шиффеля, Вейзе и других лесоводов. Н.В. Третьяковым (1927) сформулирован закон единства в строении насаждений. Хотя в ряде работ (Дыренков, 1969; Никитин, 1972; Лебков, 1973)
показано, что существуют определенные количественные вариации в распределении и некоторые отклонения от единства, однако сохраняется общее сходство в характере дифференциации деревьев на ранги, классы и т.д. и более или менее близкие пропорции в соотношении, скажем, самых крупных деревьев-лидеров (30%), средних – спутников (40 %), отстающих (30%) (Маслаков, 1984). Данное соотношение (близкое к нормальному) являясь результатом сложных взаимодействий генотипа, ценотических условий, конкуренции, эдафической мозаичности, будет наилучшим для оптимального использования ресурсов среды (солнечного света, влаги, элементов питания и пр.). Судя по распределениям, показанным на рисунке 3, у насаждений, подверженных воздействию дереворазрушающих грибов биотрофного комплекса, отсутствуют оптимальные пропорции, которые отмечает Е.Л. Маслаков. Отсутствие своевременной активной дифференциации среди отстающих деревьев, с последующим отпадом наименее приспособленных сопровождается общим ослаблением насаждения и, при наличии инфекции грибов биотрофного комплекса, распадом. В случае интенсивного воздействия Armillaria mellea sensu lato не происходит естественного и положительного для древостоя отпада только наиболее мелких деревьев. Усыхание происходит куртинами до 10 -15 деревьев. При этом погибают деревья всех классов Крафта.
Литература
- Бузыкин А.И., Пшеничникова Л.С., Суховольский В.Г. Густота и продуктивность древесных ценозов. -Новосибирск: Наука, 2002. -152 с.
- Дыренков С.А. Закономерности дифференциации деревьев по толщине и их математическая интерпретация //ЭВМ и математические методы в лесном хозяйстве. –Л.:ЛенНИИЛХ, 1969. –С. 63-75.
- Каплунов В.Я. Прогнозирование строения древостоев по диаметру // Лесоведение. -1989. -№5. -С. 68-73.
- Кузьмичев В.В., Савич Ю.Н. Влияние густоты посадки на рост сосновых культур // Лесоведение. -1979. -№6. -С. 56-63.
- Лебков В.Ф. Теория строения древостоев и ее роль в оценке продуктивности лесов //Вопросы лесоведения. –Красноярск: Наука, 1973. –С. 90-104.
- Лемешко Б.Ю. Асимптотически оптимальное группирование наблюдений в критериях согласия //Заводская лаборатория. -1998. -Т. 64. - №1. - С.56-64.
- Макаренко А.А. О свойствах рядов распределения деревьев в древостоях // Лесоведение. -1975. -№6. -С. 42-49.
- Никитин К.Е. Применение ЭВМ в лесной таксации. –М.: Урожай, 1972. – 133 с.
- Никитин К.Е., Швиденко А.З. Методы и техника обработки лесоводственной информации. -М.: Лесн. пром-ть, 1972. -272 с.
- Одум Ю. Экология. Т.1. -М.: Мир, 1986. -328 с.
- Орлов А.И. Эконометрика. – М.: Экзамен, 2002. – 576 с.
- Павлов И.Н. К методике картографирования куртинного усыхания деревьев с применением геоинформационных технологий // Лесное хозяйство – 2005. - № 4 – С.38-39.
- Павлов И.Н., Миронов А.Г. Куртинное усыхание хвойных насаждений в измененных условиях среды (вопросы диагностики) //Непрерывное экологическое образование и экологические проблемы: Сборник статей по материалам Всероссийской научно – практической конференции. Т.1. -Красноярск: СибГТУ, 2004. -С. 173-185.
- Свалов Н.Н., Свалов С.Н. О динамике рядов распределения диаметров стволов в одновозрастных сосняках // Лесоведение. -1973. - №5. -С. 58-62.
- Свирежев Ю.М., Логофет Д.О. Устойчивость биологических сообществ. -М.: Наука, 1978 . -352 с.
- Справочник по прикладной статистики. Т.1.: Пер. с англ./Под ред. Э. Ллойда, У. Ледермана, Ю.Н. Тюрина. – М.: Финансы и статистика, 1989, - 510 с.
- Справочник по прикладной статистики. Т.2.: Пер. с англ./Под ред. Э. Ллойда, У. Ледермана, С.А. Айвазяна, Ю.Н. Тюрина. – М.: Финансы и статистика, 1990. - 526 с.
- Ушанов С.В. Применение многомерных статистических методов при принятии решений. – Красноярск: СибГТУ, 2003. – 239 с.
- Феллер В. Введение в теорию вероятностей и ее приложения. Т. 2. -М.: Мир, 1984. – 738 с.
- Феллер В. Введение в теорию вероятностей и ее приложения. Т.1. - М.: Мир, 1984. - 528 с.
Shaw C. G., Kile G. A. Armillaria root disease. Agricultural handbook №691. Forest service US Department of Agriculture. – Washington D.C., 1991. –233 р.
|


