|
УДК 630*5: 630*232
К вопросу о формировании устойчивости лесных культур
И.Н. Павлов, О.А. Барабанова (Сибирский государственный технологический университет)
Для лесных культур характерно равномерное размещение деревьев на площади, исключительная одновозрастность, выровненный агрофон. Это во многом и определяет замедленный процесс дифференциации и последующий естественный отпад отстающих в росте деревьев. Слабое очищение от сучьев, а также достаточно интенсивный рост у большинства составляющих насаждение деревьев, формируют жесткую конкуренцию среди равных, что значительно увеличивает неблагоприятное взаимовлияние в биоценозе. Положительное взаимодействие резко уменьшается. Рост напряженности конкурентных отношений и является одной из главных причин пониженной устойчивости искусственно созданных фитоценозов.
Слабость гомеостатических механизмов в лесных культурах требует постоянного проведения агротехнических и лесоводственных уходов. На практике уход за культурами завершается сразу после перевода их в покрытые лесом земли, а часто и сразу после посадки. В результате - повышенная чувствительность к неблагоприятным климатическим и биогенным воздействиям и, в дальнейшем, гибель или значительное снижение продуктивности. Следовательно, нужно стремится к созданию структуры насаждения, при которой осуществляется естественная саморегуляция древостоя (при соблюдении основного правила при создании лесных культур – соответствие выбранной породы лесорастительным условиям).
Признание леса динамической системой не вызывает сомнения. Распределение деревьев по диаметру является важным показателем, характеризующим состояние древостоя. Сведения о характере динамики рядов распределения деревьев по их размерам с возрастом довольно разнородны [4-6, 11]. Вывести одну общую кривую распределения деревьев по диаметру для всех возрастов не представляется возможным [4]. На распределение деревьев по биометрическим показателям оказывает влияние большое количество факторов: возраст, условия произрастания, породный состав, полнота, характер размещения, антропогенное и биогенное воздействие, пожары, ветровалы и пр. Форма кривой распределения диаметров определяется видом и интенсивностью естественного изреживания и является его характеристикой [9].
Определенную закономерность в возрастной динамике рядов распределения диаметров стволов отмечают А. А. Макаренко [4]; Н. Н. Свалов, С. Н. Свалов [9]; В. В. Кузьмичев, Ю.Н. Савич [3]; В.Я Каплунов [2]; А. И. Бузыкин, Л. С. Пшеничникова, В. Г. Суховольский [1] и др. Лучшие закономерности были получены при наблюдениях на постоянных пробных площадях в одновозрастных древостоях, в т.ч. лесных культурах.
Для обоснованного выбора технологии создания лесных культур и дальнейшего вмешательства в процесс роста необходимо выявить долговременные закономерности, понять влияние экзогенных и эндогенных факторов в формировании структуры древостоя, его устойчивости.
Древостою, как биогеоценотической системе, свойственен свой ритм развития, который, безусловно, подвержен значительному воздействию экзогенных факторов. Одновозрастные древостои, особенно лесные культуры, имеют упрощенную структуру и легче поддаются математической интерпретации динамики строения.
В данной работе сделана попытка обобщить имеющийся собственный и опубликованный другими исследователями фактический материал, характеризующий ряды распределения лесных культур сосны обыкновенной по диаметру. Работа не претендует на всеобъемлемость выявленных закономерностей. Рассмотрен некий «идеальный» древостой, в котором строение формируется за счет доминирования внутриценотических факторов. Вмешательство экстремальных факторов (засуха, болезни, вредители, низовые пожары, ветровалы и пр.) ведет к изменению динамики показателей, характеризующих ряды распределения. Выявленные закономерности ограниченно подтверждены фактическим материалом и на данном этапе должны рассматриваться на уровне гипотезы.
Согласно современным представлениям о гомеокинезе биологические системы стремятся не к абсолютной стабилизации своего состояния, а к удержанию его в определенных пределах при непрерывном колебательном изменении параметров вокруг определенных точек равновесия [10]. Одним из механизмов поддержания стабильности в экосистеме является организация отрицательной обратной связи [7].
На разных возрастных этапах максимальный запас древостоя формируется различным количеством деревьев. Естественный отпад не только удаляет из древостоя генетически худшие экземпляры, но и формирует структуру, обеспечивающую устойчивость ценоза. В свою очередь, загущенность среди деревьев нижних ступеней толщины и, как следствие, высокая положительная асимметрия выполняют роль обратной связи, определяющей, какое количество деревьев необходимо элиминировать из древостоя.
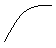
Посадка лесных культур предусматривает отбраковку некондиционных сеянцев, имеющих диаметр ниже установленного. Следовательно, искусственно отсекается левый «хвост» в распределении (формируется положительная асимметрия). В дальнейшем (1 этап продолжительностью 10-20 лет), происходит снижение асимметрии за счет двух факторов: интенсивного роста лидеров и средних деревьев (группировка в верхних ступенях толщины); отставание (без гибели) в росте неприспособленных, некачественно посаженных и пр. В итоге - отрицательная асимметрия. Высокая скорость прохождения будет свидетельствовать о быстроте формирования лесной обстановки на лесокультурной площади (рост положительного взаимодействия) и успешной дифференциации в древостое (увеличение конкуренции).
На следующем этапе (II) после смыкание крон (более длительном, до 40 -50 лет) с ростом полноты сокращаются приросты у многочисленной тонкомерной части древостоя, происходит отпад наиболее мелких и, в итоге, образуется положительная асимметрия. В процессе увеличения положительной асимметрии возрастает конкуренция и снижается текущий прирост древостоя.
На следующем (III) этапе (приспевающие - спелые насаждения) в процессе дифференциации и усилению конкурентных взаимоотношений происходит естественный отпад отстающих в росте деревьев. Катализатором отпада деревьев 5 класса Крафта может стать комплекс неблагоприятных погодных условий. Ряд деревьев 3-4 классов, используя освободившееся пространство, переходит в верхние ступени толщины; деревья-лидеры, вступая в фазу активного репродуктивного развития, снижают прирост – в итоге формируется отрицательная асимметрия. Далее при наличии второго яруса возможны дальнейшие изменения асимметрии. Динамика изменения асимметрии распределения диаметра ствола на высоте груди культур сосны обыкновенной представлена на рисунке 1. Естественно, представленный вариант не отражает всего многообразия возможных процессов динамики строения древостоя.
Для математического представления процесса в общем, идеализированном виде динамику строения некого участка лесных культур, имеющего непрерывный процесс циклического изменения в рядах распределения деревьев, можно выполнить в виде свободного затухающего во времени колебания маятника с изменяемым расстоянием от точки подвеса. Термин «маятник» широко используется в приложении к системам различной природы (физика, экология, химия, экономика). Причина – естественная цикличность природных процессов, во многом определяемых вращением Земли.
Динамика показателя асимметрии представляет наибольший интерес. Для моделирования изменения асимметрии в строении деревьев по диаметру (исходя из многочисленных литературных и собственных данных) график затухающего колебания представим в виде амплитудно-модулированной синусоиды (рис. 2). Её отличительной особенностью является различная продолжительность периодов в разные этапы жизни древостоя. Весь жизненный цикл искусственного насаждения представлен в долях от единицы. Исключено воздействие экстремальных факторов (засуха, болезни, вредители, низовые пожары, ветровалы и пр.).
Уравнение, описывающее данное колебание имеет следующий вид:
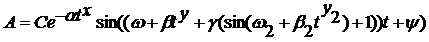 (1) (1)
Данная форма синусоиды образуется при следующих параметрах уравнения: C=0,8; a=2,2; x=1,7; w=18; w2 = 0,8; b=6,5; y=10; y2 = 0,4; b2 = 5; y=1,57; g=7
Экзогенные и внутриценотические факторы оказывают влияние на форму синусоиды (период и амплитуду) и соответственно на параметры уравнения. Вид посадочного материала, его возраст и состояние оказывают влияние на первоначальное значение асимметрии (С). Так тщательная сортировка сеянцев ведет к увеличению положительной асимметрии, а отсутствие – наоборот, к снижению.
Изменение частоты в начальный период корректируется показателем w2. При прочих равных условиях (первоначальная густота, характеристика посадочного материала) с увеличением плодородия почвы наблюдается более раннее смыкание крон и как следствие рост конкуренции и последующий отпад. Соответственно сокращается продолжительность первого периода. За изменение частоты в приспевающем и спелом возрасте отвечают b2 и y2. В перестойном возрасте основные колебания в ряду распределения происходят, в основном в процессе единичного отпада. Поэтому колебания имеют малую амплитуду, но при этом частота увеличивается (в уравнении это происходит изменением b и y).
Амплитуда колебания определяется показателями a и x. 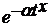 можно рассматривать как индекс напряженности конкуренции (зависит от типа размещения, свойств посадочного материала). При равномерном во времени отпаде и синхронном росте происходит быстрое затухание амплитуды. можно рассматривать как индекс напряженности конкуренции (зависит от типа размещения, свойств посадочного материала). При равномерном во времени отпаде и синхронном росте происходит быстрое затухание амплитуды.
Для верификации представленной гипотезы фактические показатели асимметрии наложены на теоретический график (рис. 3). В качестве эмпирических данных использованы результаты обмеров культур сосны обыкновенной на постоянных пробных площадях (ППП) Минусинского (1965, 1971-1972, 1981, 1991, 2002-2003 гг.), Боготольского (2002) и временных пробных площадях Большемуртинского лесхозов (2000).
Представлены насаждения, имеющие наибольший исследовательский интерес. ППП 3 заложена в насаждении, сформированном на заброшенном посевном отделении сосны обыкновенной. По первоначальной густоте (>250 тыс. шт/га) они близки к естественному возобновлению в боровых условиях произрастания.
Три пробные площади (12, 15, 22) заложены в насаждениях, нарушенных воздействием грибов биотрофного комплекса. Причина гибели деревьев установлена по комплексу диагностических признаков (Павлов, Миронов, 2004).
ППП 1, 6, 10, 13, 14, 16 несколько отличаются по первоначальной густоте, имеют средние показатели роста, устойчивое состояние и выбраны в качестве контроля. Для них характерно изменение асимметрии близкое к модельной амплитудно-модулированной синусоиде.
Для культур, имеющих негативное состояние, характерно отсутствие колебания асимметрии (ППП 12), колебание происходит только в положительной области или значительное (в два и более раза) запаздывание при переходе от положительной асимметрии к нормальному распределению на первом этапе (ППП 15, 22). Длительное нахождение большей части древостоя в загущенном состоянии в левой части распределения из за пониженной дифференциации и равномерного естественного отпада отстающих в росте деревьев негативно сказывается на состоянии древостоя в целом. Недостаток питательных веществ, влаги, освещенности испытывают большинство деревьев образующих древостой.
К сожалению, отсутствуют наблюдения за распределением деревьев по диаметру (или другим биометрическим показателям) на стационарных пробных площадях за весь жизненный цикл древостоя. Однако, даже при анализе относительно коротких периодов (30-40 лет) прослеживается выше обозначенные закономерности в колебании асимметрии. Скорость изменения асимметрии и соответственно строения древостоя во многом зависит от первоначальной густоты древостоя.
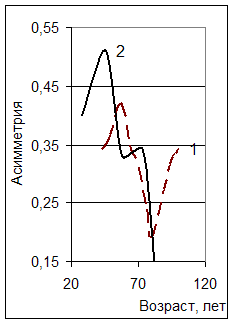
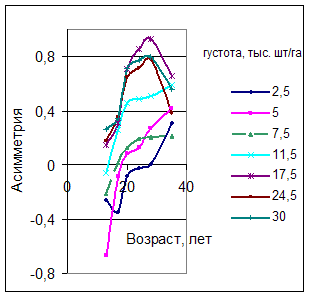
Обозначенные закономерности находят свое подтверждение в приведенных данных других авторов (рис. 4). Построенный по данным В.В Кузьмичева и Ю.Н. Савич [3] график изменения асимметрии для лесных культур разной густоты демонстрирует фрагмент амплитудно-модулированной синусоиды затухающего колебания (рис. 5). Более густые насаждения быстрее вступают в напряженные конкурентные отношения, что ведет к более ранней дифференциации древостоя.
Экстремальные факторы (низовые пожары, ветровалы, аномальные режимы увлажнения, болезни и вредители и пр.) могут оказывать значительное влияние на форму синусоиды. Возможно ускорение, замедление или изменения вектора асимметрии. Рассмотрим два момента, часто имеющие место в реальном насаждении. В первом периоде минимальных значений асимметрия достигает при отсутствии существенного отпада отстающих в росте деревьев. Отпад начинается после смыкания крон в культурах и подавления наиболее мелких особей лидерами. Ускорить процесс элиминации деревьев из нижних ступеней толщины и соответственно увеличить асимметрию способны низовые пожары, сосновая пяденица, засуха (рис. 6, синусоида «в»).
Функция, описывающая влияние указанных экстремальных факторов в динамике, является колебательным процессом асимметрии А в зависимости от времени t. Зависимость переменной А от времени в общем виде имеет вид:
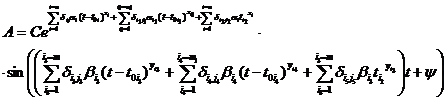 (2) (2)
где С, y – константы, определяемые из начальных условий асимметрии; е – основание натурального логарифма; n, m – необходимое число слагаемых для описания функции А, на которых производится корректировка амплитуд и частот функции А; t0 – время начала срабатывания экстремальных факторов; a, х – функции описывающие изменение амплитуды асимметрии; b, у – функции описывающие изменение частоты асимметрии; величины 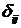 , , 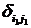 и т.д. – символы Кронекера, принимающие значения равные единице при равенстве индексов и нулю при неравенстве индексов исходя из условия непрерывности функции А и влияния экстремальных факторов; и т.д. – символы Кронекера, принимающие значения равные единице при равенстве индексов и нулю при неравенстве индексов исходя из условия непрерывности функции А и влияния экстремальных факторов; 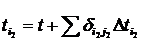 , где величины , где величины 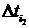 определяются из условия непрерывности функции А. определяются из условия непрерывности функции А.
Реконструкция графика асимметрии это задача синтеза, т.е. итерационная задача. Рассмотрим влияние экстремальных факторов на примере рассмотренной эмпирической формулы (1). Зависимость переменной А от времени в общем виде имеет вид:
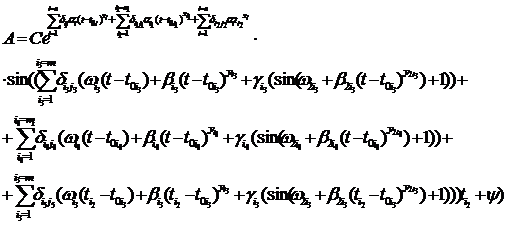 (3) (3)
Данная функция описывает срабатывание нескольких экстремальных факторов на всем периоде продолжительности жизни древостоя. Рассмотрим применение этой функции для анализа экстремальных факторов на разных отрезках времени. Допустим, что экстремальные факторы срабатывают только один раз за весь период жизни древостоя. Функция, удовлетворяющая этим условиям, имеет вид:
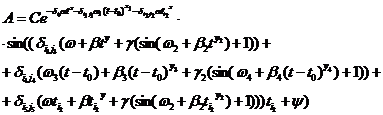 (4) (4)
где символы Кронекера 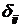 и и 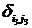 принимают значение равное единице до наступления экстремальных факторов и развитие древостоя идет аналогично формуле (1) (рис. 6, синусоида «а»), а на остальном промежутке времени принимают значение равное нулю; принимают значение равное единице до наступления экстремальных факторов и развитие древостоя идет аналогично формуле (1) (рис. 6, синусоида «а»), а на остальном промежутке времени принимают значение равное нулю; 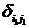 и и 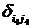 принимают значение равное единице с момента наступления экстремальных факторов до момента окончания воздействия этих факторов и развитие древостоя отклоняется от нормального (рис. 6, синусоида «в»); принимают значение равное единице с момента наступления экстремальных факторов до момента окончания воздействия этих факторов и развитие древостоя отклоняется от нормального (рис. 6, синусоида «в»); 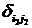 и и 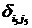 принимают значение равное единице в момент окончания воздействия экстремальных факторов и развитие древостоя продолжается аналогично синусоиды «а», но с некоторым опережением по времени. принимают значение равное единице в момент окончания воздействия экстремальных факторов и развитие древостоя продолжается аналогично синусоиды «а», но с некоторым опережением по времени.
Изменяя время вмешательства экстремальных факторов в нормальное развитие древостоя можно моделировать различные варианты динамики строения древостоя. Например, во II периоде с увеличением полноты древостоя сокращаются приросты у многочисленной тонкомерной части и образуется положительная асимметрия. Равномерное размещение посадочных мест на лесокультурной площади, выровненный агрофон, близкие характеристики посадочного материала не способствуют равномерному естественному отпаду наиболее мелких деревьев и ведут к снижению устойчивости и продуктивности древостоя. И как следствие увеличивается вероятность развития болезней и вредителей. В итоге возможно резкое снижение асимметрии (рис. 6, синусоида «с»).
Непрерывное колебательное изменение показателя асимметрии рядов распределения деревьев по диаметру можно рассматривать как необходимый атрибут формирования устойчивого насаждения. Он позволяет осуществлять поиск оптимального для данного возраста и лесорастительных условий строения. Устойчивость насаждения достигается созданием внутриценотических условий в которых положительные взаимодействия деревьев сочетаются с естественным отбором. Благодаря конкуренции и своевременной дифференциации среди слагающих древостой особей осуществляется своевременное удаление из ценоза отстающих в росте экземпляров.
Список литературы
- Бузыкин А.И., Пшеничникова Л.С., Суховольский В.Г. Густота и продуктивность древесных ценозов. Новосибирск: Наука, 2002. 152 с.
- Каплунов В.Я. Прогнозирование строения древостоев по диаметру // Лесоведение. 1989. №5. С. 68-73.
- Кузьмичев В.В., Савич Ю.Н. Влияние густоты посадки на рост сосновых культур // Лесоведение. 1979. №6. С. 56-63.
- Макаренко А.А. О свойствах рядов распределения деревьев в древостоях // Лесоведение. 1975. №6. С. 42-49.
- Моисеев В.С. Таксация молодняков. Л.: ЛТА., 1971. 343 с.
- Никитин К.Е. Лиственница на Украине. Киев, 1966. 331 с.
- Одум Ю. Экология. Т.1. М.: Мир, 1986. 328 с.
- Павлов И.Н., Миронов А.Г. Куртинное усыхание хвойных насаждений в измененных условиях среды (вопросы диагностики) //Непрерывное экологическое образование и экологические проблемы: Сборник статей по материалам Всероссийской научно – практической конференции. Т.1. Красноярск: СибГТУ, 2004. С. 173-185.
- Свалов Н.Н., Свалов С.Н. О динамике рядов распределения диаметров стволов в одновозрастных сосняках // Лесоведение. 1973. №5. С. 58-62.
- Свирежев Ю.М., Логофет Д.О. Устойчивость биологических сообществ. М.: Наука, 1978 . 352 с.
- Тябера А.П. Принципы исследований строения древостоев по толщине деревьев // Изв. вузов. Лесной журнал. 1980. №1. С. 5-8.
- Эйтинген Г.Р. Избранные труды. М.: Сельхозгиз, 1962. 250 с.
Стадии развития |
Этапы

сеянцы спелое насаждение |
Значение асимметрии |
Max
+ |
0 |
Max
- |
0 |
Max
+ |
0 |
Max
- |
0 |
Ежегодный прирост |

|
|

|
|
Период |
1 |
2 |
3 |
4 |
Рис. 1 Общие закономерности динамики строения лесных культур
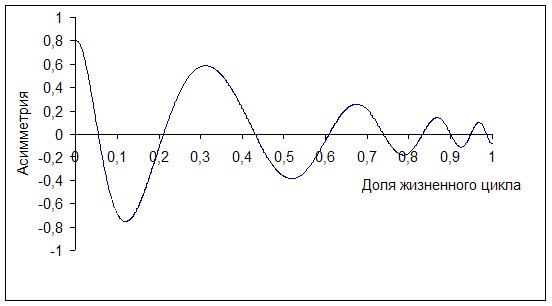
Рис. 2 Амплитудно-модулированная синусоида затухающего колебания асимметрии рядов распределения по диаметру лесных культур сосны обыкновенной
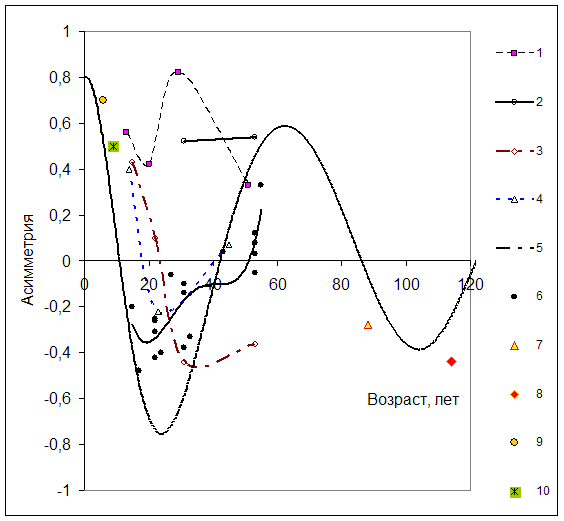
Рис. 3 Динамика показателя асимметрии рядов распределения числа стволов культур сосны по толщине (1 - ППП3; 2 - ППП12; 3 - ППП15; 4 - ППП22; 5 - амплитудо-моделированная синусоида; 6 - ППП6, 10, 13, 14, 16; 7 - ППП 1б; 8 - ППП 2б; 9 - ПП 1к; 10 - ПП 2к.
ППП 6, 10, 13, 14, 16 - культуры без признаков поражения (сглажено полиномом); 12, 15, 22 - культуры с поражением грибами биотрофного комплекса; ППП 3 – первоначальная густота >250 тыс. шт/га).
|
|
Рис. 4. Динамика показателя асимметрии рядов распределения числа стволов сосны по толщине (по Г. Р. Эйтингену [12]: 1 – естественный древостой; 2- лесные культуры) |
Рис. 5 Влияние густоты на асимметрию рядов распределения числа стволов культур сосны по толщине (по данным В.В. Кузьмичева и Ю.Н. Савич [3]) |
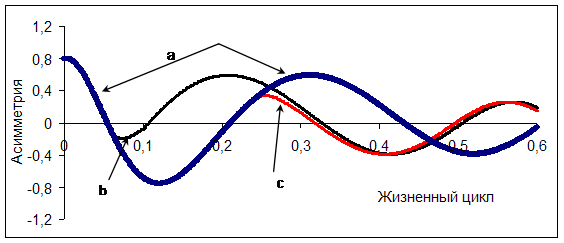
Рис. 6 Влияние экстремальных факторов на симметрию рядов распределения по диаметру (а – воздействие экстремальных внешних факторов исключено; в - низовые пожары, засуха, сосновая пяденица; с – грибы биотрофного комплекса (корневая губка, опенок обыкновенный и осенний).
Реферат
К вопросу о формировании устойчивости лесных культур
Высокая напряженность конкурентных отношений является одной из главных причин пониженной устойчивости искусственно созданных фитоценозов. На основе большого фактического материала предложена гипотеза непрерывного колебательного изменения показателя асимметрии рядов распределения деревьев по диаметру. Предложено рассматривать процесс колебания как необходимый атрибут формирования устойчивого насаждения. Он позволяет осуществлять поиск оптимального для данного возраста и лесорастительных условий строения. Устойчивость насаждения достигается созданием внутриценотических условий в которых положительное взаимодействие деревьев сочетается с естественным отбором. Благодаря конкуренции и своевременной дифференциации среди слагающих древостой особей осуществляется своевременное удаление отстающих в росте экземпляров из ценоза. Представлена математическая модель затухающего колебания (в виде амплитудно-модулированной синусоиды с различной продолжительностью периодов в разные этапы жизни древостоя). В модели предусмотрено изменение колебания показателя асимметрии под влиянием экстремальных факторов (низовые пожары, засуха, сосновая пяденица, грибы биотрофного комплекса) |


