|
УДК 630*232
Павлов И.Н., Барабанова О.А., Кузьмичев В.В., Мочалов А.А.
Анализ внутриценотических взаимодействий в лесных культурах сосны обыкновенной
На примере 53-летних лесных культур сосны обыкновенной с помощью геоинформационных технологий проведен анализ внутриценотического взаимодействия. Из всех изученных параметров наибольшее влияние на рост оказывают количество деревьев-«соседей», их размеры и расстояние до модельных экземпляров. В результат дисперсионного анализа установлено, что влияние деревьев-«соседей» объясняет почти половину дисперсии толщины ствола (42,6 %, р<0,001). В результате воздействия патогенных дереворазрушающих грибов (виды комплекса Armillariamelleasensulato; Heterobasidionannosum (Fr.) Bref.). произошло некоторое снижение установленной закономерности влияния деревьев-«соседей» на рост модельных особей по диаметру. Так если в 1981 г. связь между количеством «соседей» и диаметром модельного дерева составляет -0,59, то к 2003 году коэффициент корреляции снижается до -0,50 (р<0,001). По результатам дисперсионного анализа установлено снижение доли влияния количества расположенных рядом деревьев (до 24,9 %, р<0,001).
Конкурентные отношения между деревьями совершенно справедливо относят к ведущим факторам, оказывающим значительное влияние на рост и строение насаждений с первых лет его существования до распада (Гурцев, Николаев, 1994). Дифференциация в насаждении происходит в результате различий в генотипе, эдафической мозаичности, воздействия вредителей и болезней и множества случайных факторов (качество посадки и ухода, давление со стороны естественного возобновления лиственных). Слабая дифференциация наблюдается в случае однородности деревьев по наследственным свойствам. В результате происходит «эффект группового угнетения» (Погребняк, 1968; Романовский, 2002). Описаны случаи, когда первоначально загущенные лесные культуры к возрасту спелости превращаются в редины (Рябоконь, 1979). В.В. Кузьмичев и О.П. Секретенко (1998) отмечают ускоренный рост деревьев, вокруг которых в результате случайного отпада в раннем возрасте образовалось малозаселенное пространство, а в тех местах, где сохранилась большая часть посаженных экземпляров, все они, конкурируя между собой, отстают в росте. Однако у этой закономерности, не вызывающей возражения, в культурах достаточно часто имеют место исключения (чаще чем в древостоях естественного происхождения). В данной работе сделана попытка выделить долю влияния ценотических факторов на рост насаждения.
Условия, которые вызывают дифференциацию деревьев в естественном древостое, очень сильно снижают свое влияние в культурах. Как правило, все деревья здесь одного возраста, находятся в идентичных почвенно- климатических условиях. Различия в наследственной энергии роста сглажены их сортировкой. Регулярная обработка почвы устраняет конкуренцию с сорной растительностью. Вместо хаотического размещения – идеально равномерное распределение по площади с одинаковым шагом посадки. Благодаря этому условия роста более выровнены, меньше выражена изменчивость высот, диаметров и других таксационных показателей. Казалось бы, что идеально подобранные условия должны улучшить качество древостоя, но борьба за существование ведется между «одинаково сильными» деревьями, что приводит к большему отпаду и неоправданности затрат на выращивание.
При помощи геоинформационной программы ArcMap было представлено размещение деревьев сосны обыкновенной на постоянных пробных площадях. Программа позволяет не только хранить картографическую информацию, автоматизировать расчеты таксационных показателей, но и осуществлять моделирование процессов роста и развития древостоев. Необходимым этапом познания закономерностей взаимоотношения организмов со средой обитания является изучение пространственной структуры популяции. Ценотическая устойчивость и, следовательно, высокая производительность лесных культур определяется оптимальным размещением деревьев различных категорий крупности. Исследование динамики формирования пространственной структуры производного насаждения, т.е. характера размещения деревьев разных пород и возраста позволяет выявить механизмы формирования биогенетической, морфологической и функциональной структуры древостоев и законов их биологической устойчивости. Анализ размещения древесных особей в насаждении дает неизмеримо большую информацию о природе изучаемого леса и ее подлинном разнообразии, чем метод суммарного описания типа леса с интерпретацией сведений о его растительности в виде различных усредненных индексов (средней высоты и диаметра, состава и полноты древостоя) (Емшанов, 1999). Особый интерес представляет динамика пространственных структур в процессе организации насаждения, другими словами, когда древесные растения начинают взаимодействовать не только с окружающей средой, но и друг с другом. Установление математических закономерностей взаимодействия значительно упрощается с применением геоинформационных технологий.
Пример отображения пробной площади № 12 приведен на рисунке 1. Результаты инвентаризации за период 1981 – 2003 гг. представлены слоями.
Для установления закономерностей роста модельных деревьев в биогруппах по диаметру, в определенном радиусе (максимальный диаметр кроны на ППП на год обследования) от модельных деревьев были количественно учтены все деревья (n 81, n 03 – соответственно в 1981 и 2003 гг.), а также рассчитан ряд коэффициентов: k 81, k 03 –отношения диаметра дерева «соседа» (на высоте 1,3 м) к расстоянию до модельного дерева (сумма всех «соседей»); kb 81, kb 03 – сумма ранговых коэффициентов деревьев-«соседей». Результаты корреляционного анализа приведены на рисунке 2.
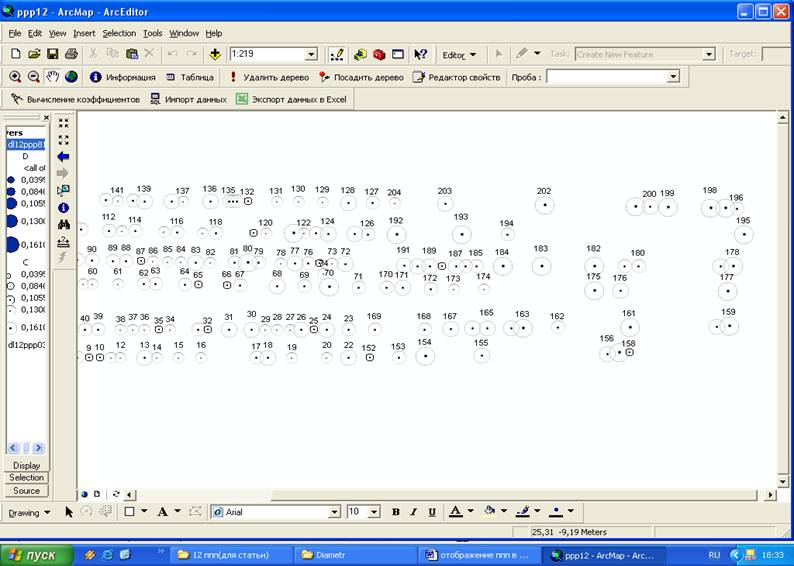
Рисунок 1- Фрагмент размещения деревьев в программе ArcMap (ППП 12, 2003 г.)
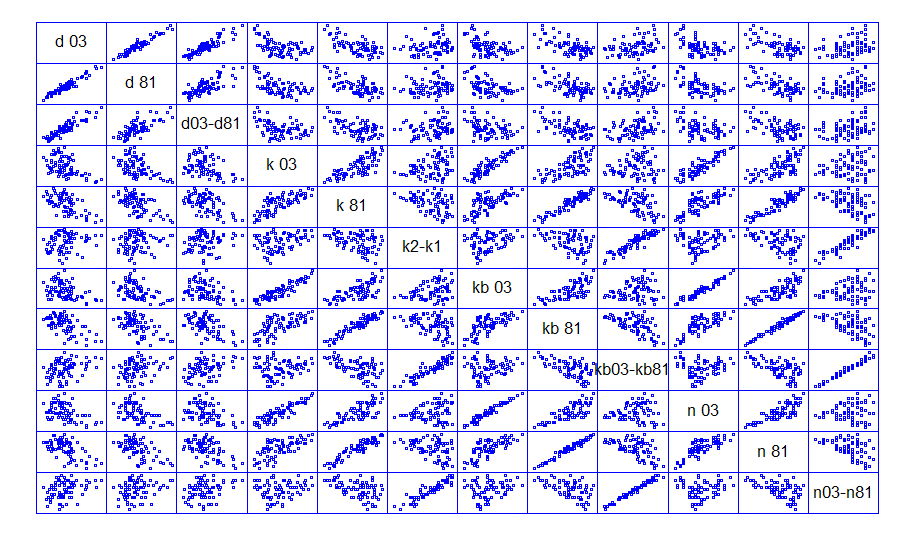
Рисунок 2 - Корреляционная матрица рассеяния (d 81, d 03 – диаметры деревьев в 1981 и 2003 гг. соответственно; n 81, n 03 – количество деревьев «соседей» в 1981 и 2003 гг. соответственно; k 81, k 03, kb 81, kb 03 – коэффициенты влияния)
Таблица 1 Корреляционная матрица взаимовлияний
Показатель |
Коэффициент корреляции, уровень вероятности |
d 81 |
d 03 |
d03-d81 |
k 81 |
k 03 |
k2-k1 |
kb 81 |
kb 03 |
kb2-kb1 |
n 81 |
n 03 |
n03-n81 |
d 81 |
r |
1 |
|
|
|
|
|
|
|
|
|
|
|
d 03 |
r |
0,96 |
1 |
|
|
|
|
|
|
|
|
|
|
p |
0,00 |
|
|
|
|
|
|
|
|
|
|
d03-d81 |
r |
0,75 |
0,90 |
1 |
|
|
|
|
|
|
|
|
|
p |
0,00 |
0,00 |
|
|
|
|
|
|
|
|
|
k 81 |
r |
-0,62 |
-0,64 |
-0,56 |
1 |
|
|
|
|
|
|
|
|
p |
0,00 |
0,00 |
0,00 |
|
|
|
|
|
|
|
|
k 03 |
r |
-0,53 |
-0,52 |
-0,42 |
0,74 |
1 |
|
|
|
|
|
|
|
p |
0,00 |
0,00 |
0,00 |
0,00 |
|
|
|
|
|
|
|
k2-k1 |
r |
0,13 |
0,17 |
0,21 |
-0,36 |
0,35 |
1 |
|
|
|
|
|
|
p |
0,26 |
0,13 |
0,07 |
0,00 |
0,00 |
|
|
|
|
|
|
kb 81 |
r |
-0,58 |
-0,62 |
-0,57 |
0,94 |
0,65 |
0,41 |
1 |
|
|
|
|
|
p |
0,00 |
0,00 |
0,00 |
0,00 |
0,00 |
0,00 |
|
|
|
|
|
kb 03 |
r |
-0,46 |
-0,44 |
-0,35 |
0,67 |
0,91 |
0,32 |
0,69 |
1 |
|
|
|
|
p |
0,00 |
0,00 |
0,00 |
0,00 |
0,00 |
0,00 |
0,00 |
|
|
|
|
kb2-kb1 |
r |
0,29 |
0,35 |
0,38 |
-0,53 |
0,12 |
0,92 |
-0,60 |
0,17 |
1 |
|
|
|
p |
0,01 |
0,00 |
0,00 |
0,00 |
0,29 |
0,00 |
0,00 |
0,15 |
|
|
|
n 81 |
r |
-0,59 |
-0,61 |
-0,54 |
0,93 |
0,65 |
-0,39 |
0,98 |
0,70 |
-0,56 |
1 |
|
|
p |
0,00 |
0,00 |
0,00 |
0,00 |
0,00 |
0,00 |
0,00 |
0,00 |
0,00 |
|
|
n 03 |
r |
-0,51 |
-0,50 |
-0,39 |
0,73 |
0,89 |
0,22 |
0,75 |
0,97 |
0,06 |
0,79 |
1 |
|
p |
0,00 |
0,00 |
0,00 |
0,00 |
0,00 |
0,05 |
0,00 |
0,00 |
0,62 |
0,00 |
|
n03-n81 |
r |
0,25 |
0,30 |
0,33 |
-0,48 |
0,18 |
0,92 |
-0,55 |
0,21 |
0,98 |
-0,52 |
0,12 |
1 |
p |
0,03 |
0,01 |
0,00 |
0,00 |
0,11 |
0,00 |
0,00 |
0,06 |
0,00 |
0,00 |
0,31 |
Для оценки взаимовлияния деревьев в ценозе проведен дисперсионный анализ. Из всех изученных параметров наибольшее влияние на рост оказывает количество деревьев-«соседей» (N) , их размеры и расстояние до модельных особей. Рассчитаны (К, Кв) коэффициенты, учитывающие не только их количество, но также и диаметр деревьев. В расчет включены особи, расположенные в определенном, зависящем от максимального размера кроны в насаждении, радиусе от модельного (табл.1). В связи с тем, что различие между коэффициентами корреляции не существенно и установление простого количества соседних деревьев значительно упрощает процедуру анализа, за основу взято значение N. В качестве зависимой величины принят диаметр на высоте 1,3 м. Для проведения дисперсионного анализа данные распределены в блоки: по количеству деревьев-соседей (4 блока); по повторностям (индивидуальное значение).
В таблице 2 показаны итоговые данные дисперсионного анализа. Значимым является линейный эффект влияния соседних деревьев. Данные позволяют оценить степень влияния основных факторов на показатель-отклик (табл. 2).
Таблица 2
Дисперсионный анализ влияния (1981 г.)
Категория изменчивости |
Число степеней свободы |
Сумма квадратов |
Среднее |
F |
р-уровень |
Влияние: |
|
|
|
|
|
Количество «соседей» |
3 |
257,08 |
85,69 |
17,550 |
0,000000 |
По повторностям |
24 |
102,89 |
4,29 |
0,878 |
0,626952 |
Остатки |
50 |
244,14 |
4,88 |
|
|
Таблица 2
Оценка степени влияния факторов (1981 г.)
Факторы |
Сумма квадратов |
Степень влияния (Кj), % |
р-уровень |
Количество «соседей» |
257,08 |
42,6 |
<0,001 |
По повторностям |
102,89 |
17,0 |
>0,05 |
Установленные факторы |
359,97 |
59,6 |
|
Случайные факторы и ошибки |
244,14 |
40,4 |
|
Все факторы |
604,11 |
100,0 |
|
Из результатов анализа следует, что влияние деревьев-«соседей» объясняет почти половину дисперсии толщины ствола (42,6 %, с уровнем значимости <0,001). На случайные факторы и ошибки приходится 40,4 % дисперсии. Причина столь высокого значения заключается во влиянии множества факторов на рост культур по диаметру. Влияние повторностей оказалось несущественно. Следовательно, в них не содержится какой-либо закономерности, что позволяет их дисперсию отнести на не установленные факторы. Таким образом, 57,4 % дисперсии может быть отнесено на влияние генотипа, почвенной мозаичности, вредителей и болезней, ошибки при посадке, а также при проведении измерений.
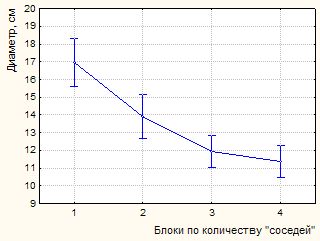
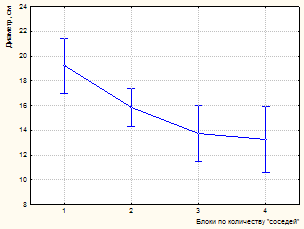
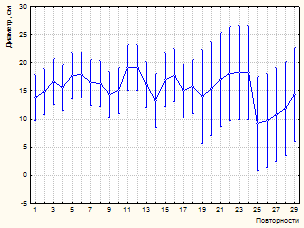
К 2003 году на пробной площади и в насаждении в целом произошел значительный отпад. Усыхание имеет ярко выраженную куртинность. Погибают деревья без явных признаков угнетения. По ряду диагностических признаков установлено воздействие дереворазрушающих грибов биотрофного комплекса (виды комплекса Armillaria mellea sensu lato; Heterobasidion annosum (Fr.) Bref.). Это явились причиной некоторого снижения установленной закономерности влияния деревьев-«соседей» на рост модельных особей по диаметру. Так, если в 1981 г. связь между количеством «соседей» и диаметром модельного дерева составляет -0,59, то к 2003 году коэффициент корреляции снижается до -0,50 (р<0,001). По результатам дисперсионного анализа (табл. 3, 4) установлено снижение доли влияния количества рядом расположенных деревьев (до 24,9 %, р<0,001). На рисунке 3 показано влияние изученных факторов на изменение диаметра модельных деревьев.
Таблица 3
Дисперсионный анализ влияния (2003 г.)
Категория изменчивости |
Число степеней свободы |
Сумма квадратов |
Среднее |
F |
р-уровень |
Влияние: |
|
|
|
|
|
Количество «соседей» |
3 |
359,126 |
119,709 |
7,2657 |
0,0004 |
По повторностям |
28 |
322,640 |
11,523 |
0,6994 |
0,8420 |
Остатки |
46 |
757,891 |
16,476 |
|
|
Таблица 4
Оценка степени влияния факторов (2003 г.)
Факторы |
Сумма квадратов |
Степень влияния (Кj), % |
р-уровень |
Количество «соседей» |
359,126 |
24,9 |
<0,001 |
По повторностям |
322,640 |
22,4 |
>0,05 |
Установленные факторы |
681,766 |
47,4 |
|
Случайные факторы и ошибки |
757,891 |
52,6 |
|
Все факторы |
1439,657 |
100,0 |
|
Рисунок 3 – Влияние факторов (количество «соседей», повторности) на изменение диаметра модельных деревьев (А, В – инвентаризация 1981 г., С, D –2003 г.)
Литература:
- Кузьмичев В.В., Секретенко О.П. Горизонтальная структура и динамика состава смешанных культур лесообразователей // Реконструкция гомеостаза: Материалы IX Международного симпозиума, 1998.–Т 2. – С. 89-95.
- Погребняк П.С. Общее лесоводство. –М.: Колос, 1968. - 440 с.
- Романовский М.Г. Продуктивность, устойчивость и биоразнообразие равнинных лесов европейской России. –М.: МГУЛ, 2002. – 97 с.
- Рябоконь А.П. Определение биологического оптимума густоты сосновых древостоев в условиях свежей субори //Лесоведение. -1979. -№ 3. –С. 16-23.
|



 А)
А) 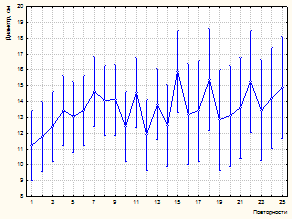
 D)
D)