|
"Хвойные бореальной зоны" 2008г.,№3-4, с.
284-289
ОСОБЕННОСТИ РОСТА И МЕТАБОЛИЗМА ЛИСТВЕННИЦЫ СИБИРСКОЙ В УСЛОВИЯХ ФИТОЦЕНОТИЧЕСКОГО СТРЕССА
Плаксина И.В.
Институт леса им. В.Н.Сукачева СО РАН
660036 Красноярск, Академгородок, 50; е-mail: biochem@ksc.krasn.ru
Исследовали состояние 17-летних культур лиственницы сибирской (LarixsibiricaLedeb.), заложенных на серых лесных почвах в подзоне южной тайги Средней Сибири с начальной густотой 0,5, 10 и 128 тыс. экз./га. Комплексный морфолого-анатомический анализ деревьев показал, что фитоценотический стресс вызывает уменьшение размеров и массы структурных частей деревьев, ширины годичного кольца и приводит к изменению соотношения поздней и ранней древесины. При этом происходит общее снижение содержания запасных веществ (белков, жиров, углеводов) в органах и тканях деревьев. Выявлены специфические изменения метаболизма, которые могут служить биохимическими индикаторами фитоценотического стресса.
Ключевые слова: лиственница сибирская, густота посадки, ксилогенез, запасные вещества
The comparative investigation of 17-years stands of Larix sibirica Ledeb. with different initial densities (0,5, 10, 128 th./h) located in south taiga zone of Central Siberia was carried out. It was found that phytocenotic stress impact induced the diminishing of mass and length of trees structural parts, width of annual ring wood and lead to increase of late tracheids number in annual wood ring. In the high density plantation the main storage componunds content (protein nitrogen, oil and starch) in organs and tissues of trees was decreased. Some of discovered specific metabolic reactions can be used like an indicators of phytocenotic stress.
Key words: Larix sibirica, stand density, xylogenesis, storage compounds
Введение
Продуктивность лесных насаждений в Сибири, где сосредоточена половина запасов древесины России, зависит от действия неблагоприятных факторов среды. На формирование древостоев помимо абиотических и антропогенных стрессов оказывает влияние фитоценотический стресс. Фитоценотический стресс – негативное воздействие конкурентных взаимоотношений между отдельными особями в древостое, приводящее к снижению интенсивности роста деревьев в насаждениях высокой густоты.
При фитоценотическом стрессе растения конкурируют за ресурсы, к которым относятся свет, вода и элементы минерального питания. Конечная густота древостоев, следовательно, их продуктивность определяется уровнем обеспеченности жизненными ресурсами конкретного местообитания. Увеличивающийся недостаток ресурсов в загущенном древостое может приводить к элиминации значительной части деревьев древостоя (Сукачев, 1953; Карпов, 1969).
В лесных растительных сообществах вследствие большой продолжительности жизни их основных эдификаторов – древесных растений – обостряются конкурентные отношения как между представителями разных поколений, так и в пределах одного поколения. Изменения морфометрических и физиологических показателей угнетенного подроста изучены достаточно подробно (Морозов, 1912; Злобин, 1970; Татаринов, 1972; Судачкова, Балмаева, 1974). Также много работ посвящено исследованию роста деревьев после проведения рубок ухода (Давыдов, Беляев, 1940; Тимофеев, 1950; Георгиевский, 1957; Жуков, Савельев, 1958). Конкуренция между деревьями проявляется в различиях темпов роста и интенсивности физиологических процессов и метаболизма (Судачкова, 1977; Судачкова и др., 1967). Однако, представление о механизмах влияния конкуренции на рост древесных растений окончательно еще не сформировалось. Для этого необходимо проводить комплексное изучение процессов роста и метаболизма подверженных стрессу деревьев, по сравнению со свободнорастущими в тех же условиях.
Данная работа посвящена малоизученному в настоящее время вопросу изменения роста и метаболизма деревьев лиственницы в чистом одновозрастном насаждении под влиянием только фитоценотического стресса.
Экспериментальная часть
Объектами исследования служили 19-летние деревья лиственницы сибирской (Larix sibirica Ledeb.). Культуры были созданы в 1982 году в подзоне южной тайги Средней Сибири в Большемуртинском лесхозе Красноярского края на серых лесных почвах в однородных лесорастительных условиях плакора с начальной густотой 0,5, 10 и 128 тыс. экз./га. Для посадки использовали двухлетние сеянцы. Площадь под опытные посадки, на которой в прошлом произрастали темнохвойные древостои с участием сосны и лиственницы, долгое время находилась в сельскохозяйственном обороте после освоения приенисейской территории первопоселенцами. Этот длительный эксперимент рассчитан на получение информации о структуре, росте, продуктивности, сроках вступления в репродуктивный период, биометрических показателях особей и сообществ, форме стволов и других зависимых от густоты характеристик (Бузыкин и др., 2002). Некоторые биометрические характеристики деревьев приведены в таблице 1 (Бузыкин, Пшеничникова, 2001).
Таблица 1 – Средние биометрические характеристики 19-летних деревьев лиственницы сибирской из фитоценозов различной густоты
Исходная густота, тыс.экз./га |
Текущая густота, тыс.экз./га |
Высота,
м |
Диаметр,
см |
Длина кроны,
м |
Относитель-
ная высота*, % |
0,5 |
0,2 |
5,2 |
7,3 |
5,2 |
71 |
10 |
6,7 |
6,1 |
6,9 |
3,0 |
88 |
128 |
34,3 |
5,6 |
4,2 |
1,8 |
133 |
Примечание: *- относительная высота – отношение высоты к диаметру
В конце вегетации и в начале июля следующего года с 12 средних модельных деревьев лиственницы отбирали образцы хвои и луба годичных боковых побегов, ствола и скелетных корней для биохимических анализов. Определяли следующие биометрические параметры: длина однолетних терминальных побегов и хвои брахибластов, масса хвои, масса лубяной ткани на единицу камбиальной поверхности ствола и корней. Масса абсолютно сухой лубяной ткани ствола и корней на единицу камбиальной поверхности определялась путем взвешивания высечек луба площадью 5 см2 с каждого дерева с последующим определением влажности. Для анатомических исследований брали керны древесины ствола возрастным буравом на высоте 50 см от поверхности почвы. Параметры деревьев с участка густоты посадки 0,5 тыс. экз./га служили контролем.
Определение общего и белкового азота проводили по стандартному методу Къельдаля (Плешков, 1968). Низкомолекулярные углеводы определяли по обесцвечиванию жидкости Фелинга (Вознесенский и др., 1962). Крахмал извлекали хлорной кислотой и определяли колориметрически (Humphreys, Kelly, 1961). Свободные аминокислоты экстрагировали и очищали по стандартной методике (Плешков, 1968). Количественное определение аминокислот проводили на автоматическом аминокислотном анализаторе ААА-339М. Для определения триацилглицеридов проводили экстракцию исследуемого материала смесью хлороформ : этиловый спирт в соотношении 2 : 1 в аппарате Сокслета, подвергали омылению и содержание жирных кислот определяли колориметрически (Смолистые вещества …, 1968; Ермаков и др., 1972: Methods …, 1984). В очищенном суммарном экстракте определяли фенольные соединения при помощи реактива Фолина-Дениса по стандартной методике (Ермаков и др., 1972).
Обсуждение результатов
По мере увеличения густоты произрастания у деревьев наблюдается значительное уменьшение средних диаметров, при этом средние высоты меняются незначительно, вследствие чего резко возрастает относительная высота (см. табл. 1). Такая тенденция к преимущественному росту в высоту давно была отмечена многими лесоводами и свидетельствует о том, что рост в высоту в условиях фитоценотического стресса доминирует над ксилогенезом, являясь фактором, обеспечивающим выживание. Отпад на участке с исходной густотой 10 тыс. экз./га составил 40 %, на участке 128 тыс. экз/га – 27 %.
Измеренные морфометрические параметры 19-летних деревьев лиственницы сибирской осенью и следующим летом приведены в таблице 2.
Осенью длина годичных боковых побегов под влиянием стресса на участке с густотой 128 тыс. экз./га уменьшилась на 30 %, по сравнению с контрольными деревьями. Следующим летом на участке 10 тыс. экз./га длина годичных боковых побегов была меньше в 5 раз. У деревьев с участка 128 тыс. экз./га годичных боковых побегов не было обнаружено. Длина хвои брахибластов летом значимо снизилась у деревьев с участков густоты посадки 10 и 128 тыс.экз./га, по сравнению с деревьями на контрольном участке, тогда как осенью наблюдались ее незначительные изменения. Масса хвои осенью уменьшалась в 1,2 и 2 раза на загущенных участках, по сравнению с контрольным, летом – уже в 1,5 и 3 раза.
Таблица 2 – Биометрические показатели деревьев лиственницы сибирской в фитоценозах различной густоты осенью в конце вегетации и следующим летом в середине июля
Показатели |
Осенью |
Летом |
Густота посадок, тыс. экз./га |
0,5 |
10 |
128 |
0,5 |
10 |
128 |
Длина: побегов, см |
13,6±0,7 |
11,4±1,6 |
9,9±0,9 |
12,5±0,4 |
2,3±0,1 |
0 |
хвои, мм |
31,5±0,5 |
31,7±0,5 |
31,3±0,6 |
34,3±0,9 |
26,9±0,7 |
23,6±0,7 |
Масса, г: 100 хвоинок |
0,2±0,02 |
0,12±0,01 |
0,08±0,01 |
0,18±0,01 |
0,12±0,01 |
0,07±0,01 |
1см2 луба ствола |
1,20±0,04 |
0,4±0,02 |
0,3±0,01 |
-* |
- |
- |
1 см2 луба корней |
0,64±0,05 |
0,63±0,04 |
0,40±0,03 |
- |
- |
- |
Примечание: *- измерения не проводили
Фитоценотический стресс вызывает ослабление нисходящего транспорта, о чем свидетельствует снижение с увеличением густоты посадок массы луба, приходящейся на единицу камбиальной поверхности ствола и корней. Этот показатель позволяет сравнивать реальную обеспеченность процесса ксилогенеза азотом и запасными веществами углеводной и липидной природы при разных густотах и служит косвенным подтверждением снижения запасающей функции луба под влиянием фитоценотического стресса. Масса лубяной ткани на единицу поверхности древесины ствола сократилась на участках с густотой 10 и 128 тыс. экз/га, по сравнению с контрольным участком, в среднем в 3 раза. Масса лубяной ткани корней на участке с густотой 10 тыс. экз/га отличалась от контроля незначительно, а на участке 128 тыс. экз/га была меньше почти в 2 раза.
Таким образом, фитоценотический стресс вызывает уменьшение размеров и массы структурных частей деревьев, в том числе ассимиляционного
аппарата.
Изменение параметров последнего годичного кольца древесины 19-летних деревьев лиственницы сибирской при разной густоте посадке приведено в таблице 3.
Число трахеид в радиальном ряду годичного кольца у деревьев при увеличении густоты до 10 и 128 тыс. экз/га уменьшается в 5 и 20 раз соответственно, по сравнению со свободнорастущими деревьями. При этом снижаются диаметр как ранних, так и поздних трахеид, но для ранних это снижение не превышает 22 %, а для поздних составляет 138 %. Толщина клеточной стенки трахеид значимо снижается только для поздних трахеид (Р?0.001) у деревьев с густоты посадки 128 тыс. экз./га. Характеристики поздней древесины, следовательно, изменяются в большей степени, чем ранней, при этом отмечается значительное увеличение доли поздних трахеид в годичном слое при возрастании интенсивности стресса.
Таблица 3 – Средние характеристики последнего годичного кольца древесины ствола 19-летних деревьев лиственницы сибирской при разной густоте посадки
Густота тыс. экз./га |
Число трахеид в годичном кольце |
Доля поздней древесины
% |
Диаметр трахеид древесины, мкм |
Толщина клеточной стенки трахеид древесины, мкм |
ранней |
поздней |
ранней |
поздней |
0,5 |
144±10 |
31,6±3,3 |
44,9±1,4 |
27,6±0,5 |
5,2±0,1 |
7,9±0,2 |
10 |
31±5 |
57,7±3,2 |
42,0±1,2 |
21,1±0,4 |
4,3±0,1 |
7,1±0,2 |
128 |
7±4 |
58,2±1,3 |
35,2±0,8 |
20,1±0,4 |
4,1±0,1 |
5,8±0,1 |
Анализ ширины годичных колец по годам позволяет установить возраст, в котором действие фитоценотического стресса становится очевидным, т.е. с какого времени рост по диаметру снижается в пользу роста в высоту. Как видно на рисунке 1, до 7-летнего возраста число трахеид в радиальном ряду годичного кольца лиственницы сибирской при всех густотах нарастало синхронно. У деревьев на участке с начальной густотой 128 тыс. экз./га этот показатель начинает резко снижаться по достижении 8-летнего возраста, а на участке с густотой 10 тыс. экз./га – с 9 лет.
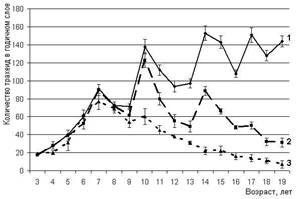
Рисунок 1 – Изменение числа трахеид в годичном слое древесины ствола лиственницы сибирской при разной густоте посадки, тыс. экз./га: 1 – 0,5, 2 – 10, 3 – 128
При сопоставлении данных по изменению количества трахеид в годичном кольце древесины с метеорологическими данными было отмечено, что в посадках с исходной густотой 10 тыс. экз./га у деревьев присутствует некоторая динамика ширины годичного кольца, синхронная с наблюдаемой у свободнорастущих деревьев в ответ на изменение погодных условий. У деревьев с участка исходной густоты в 128 тыс. экз./га наблюдается постепенное снижение числа трахеид в радиальном ряду годичного кольца древесины деревьев, начиная с 7-8-летнего возраста, что свидетельствует о снижении реакции на погодные изменения.
Для определения обеспечения процесса ксилогенеза необходимыми метаболитами в органах и тканях деревьев исследовали содержание основных запасных веществ хвойных: жиров, белков, крахмала, а также низкомолекулярных форм углеводов. Также изучали содержание свободных аминокислот, важных с точки зрения синтеза белков и количество водорастворимых фенольных соединений, выполняющих многообразные важные функции в растениях, в том числе использование их в качестве запасного энергетического материала.
Было выявлено, что при увеличении густоты посадки у деревьев лиственницы сибирской происходит увеличение концентрации общего и белкового азота в тканях. Подобные результаты были получены ранее при исследованиях подроста под материнским пологом (Судачкова и др., 1967) и разногустотных посадок сосны (Судницына, 1967). Однако, мы задались вопросом сравнить не только концентрацию азота в одном грамме сухого вещества ткани, но и проанализировать изменение содержания азота в отдельных структурах деревьев. Такой подход, по-видимому, является наиболее оптимальным, поскольку отражает реальную обеспеченность азотом, например, одной хвоинки или отдельно взятого дерева. При сравнении отдельных структур угнетенных и свободнорастущих деревьев было обнаружено значительное уменьшение содержания азота (рис. 2).
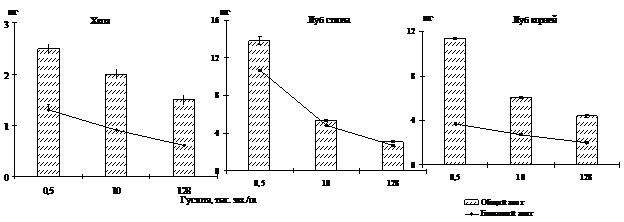
Рисунок 2 – Содержание общего и белкового азота в хвое (100 шт) и в одном см2 луба, приходящегося на один см3 камбиальной поверхности ствола и корней лиственницы сибирской в фитоценозах различной густоты
В расчете на 100 хвоинок содержание как общего, так и белкового азота при увеличении густоты посадки снижается на участках максимальной густоты, по сравнению с контрольными деревьями, более чем в 1,5-2 раза. В лубе различия еще значительнее: на единицу камбиальной поверхности ствола и корней содержание азота в варианте с максимальной густотой посадки снижается в 4 раза. Также было выявлено снижение содержания азота в расчете на 100 хвоинок и на единицу камбиальной поверхности ствола деревьев и следующим летом.
Содержание свободных аминокислот в тканях последовательно снижается с увеличением густоты во всех тканях, кроме луба ствола и корней (рис. 3а). Это может быть связано с соотношением скоростей оттока аминокислот из ствола и синтеза белка в корнях. В хвое общее количество аминокислот снизилось сократилась на участках с густотой 10 и 128 тыс. экз/га, по сравнению с контрольным участком, в среднем в 2 раза. При пересчете на 100 хвоинок эта разница оказалась еще значительнее - в 3 и 5 раз соответственно (рис. 3б).
Количество свободных аминокислот летом у деревьев с участков густоты посадки 10 и 128 тыс. экз./га снижалось во всех тканях, по сравнению со свободнорастущими деревьями. Весьма значительное уменьшение содержания свободных аминокислот было отмечено в лубе годичных боковых побегов у деревьев с участка максимальной густоты посадки, по сравнению с деревьями контрольного участка– в 16 раз.
Содержание углеводов в зависимости от густоты произрастания менялось неоднозначно, за исключением крахмала (рис. 4). Так, концентрация моносахаров при увеличении густоты посадки в лубе годичного побега и ствола уменьшалась, а в лубе корней, наоборот, увеличивалась. В хвое при очень больших концентрациях моносахаров, олигосахаров не было обнаружено. В лубе корней содержание олигосхаров уменьшается, а в лубе ствола, наоборот, резко увеличивается при возрастании фитоценотического стресса. Таким образом, можно
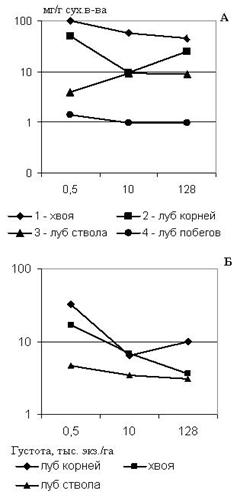
Рисунок 3 – Изменение суммарного количества свободных аминокислот при фитоценотическом стрессе: концентрация (А) в тканях (мг/г сух.в-ва) и содержание (Б) в хвое (100 шт) и в одном см2 луба, приходящегося на один см3 камбиальной поверхности ствола и корней (мг) лиственницы сибирской
отметить увеличение концентрации олигосахаров в лубе ствола угнетенных деревьев, за счет чего и наблюдается большее суммарное количество углеводов в этой ткани при возрастании густоты посадки деревьев. Тем не менее, суммарное количество неструктурных углеводов в хвое, лубе годичных боковых побегов и лубе корней в целом снижается при увеличении густоты посадки у деревьев лиственницы.
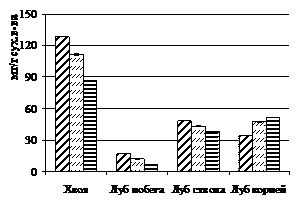
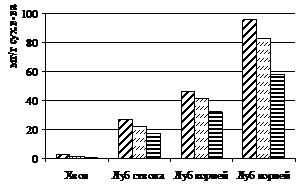
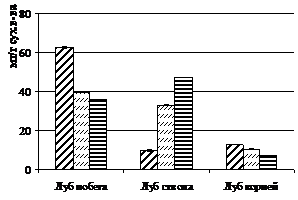
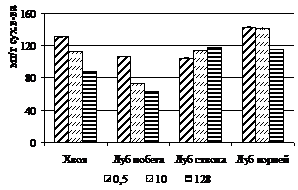
Рисунок 4 – Распределение неструктурных углеводов по органам и тканям лиственницы сибирской в посадках различной густоты: 1 - моносахаров, 2 – олигосахаров, 3 – крахмала, 4 – суммарного количества углеводов
Среди изученных групп углеводов только изменение уровня крахмала достоверно и однозначно снижается при фитоценотическом стрессе. Его концентрация сократилась в среднем в 1,5 раза в тканях наиболее угнетенных деревьев лиственницы. При этом можно проследить процесс оттока углеводов из осевых органов и накопление крахмала в корнях, что является характерным процессом для хвойных осенью (Судачкова, 1977). Содержание крахмала в расчете на 100 хвоинок и на 1см2 луба с увеличением густоты также закономерно снижается. При густоте посадки 128 тыс. экз./га запасы крахмала снизились в тканях в 3-5 раз, по сравнению со свободнорастущими деревьями.
Летом в тканях угнетенных деревьев лиственницы сибирской обнаружено достоверное снижение моносахаров и крахмала, по сравнению со свободнорастущими деревьями. Содержание олигосахаров во всех тканях с участка густоты посадки 10 тыс. экз./га увеличивается, по сравнению со свободнорастущими деревьями. У деревьев с участка густоты 128 тыс. экз./га, однако, содержание олигосахаров резко снижается, не достигая даже уровня, наблюдаемого в тканях свободнорастущих деревьев. Видимо, это связано с тем, что эти деревья находятся в фазе крайнего истощения, и в их тканях уменьшение крахмала связано прежде всего с тем, что моносахара, по всей видимости, идут на обеспечение процессов поддержания организма, а не роста, как у менее угнетенных деревьев с участка густоты посадки 10 тыс. экз./га. Общее количество неструктурных углеводов во всех тканях снижается при увеличении густоты посадки деревьев.
Жиры наряду с углеводами являются важнейшим энергетическим резервом древесных растений. Помимо основной, запасающей, функции также важна роль жиров как защитных веществ в условиях отрицательных зимних температур. Нами было обнаружено, что под влиянием фитоценотического стресса запасы жиров в тканях деревьев лиственницы сибирской значительно уменьшаются (рис. 5а). Еще более значительным сокращение содержания жиров оказывается в пересчете на отдельные структуры деревьев.
В хвое количество жиров в 100 хвоинках деревьев с участка густоты посадки 128 тыс. экз./га уменьшается в 10 раз, по сравнению со свободнорастущими деревьями, в лубе ствола – в 7 раз, а в лубе корней это снижение значимо, но в меньшей степени.
В жестких климатических условиях Сибири жиры (триацилглицериды) у хвойных являются основной формой запасных соединений. Наибольшей способностью к запасанию жиров среди тканей вегетативных органов отличается лубяная ткань (Судачкова, 1977). При этом наибольшее количество триацилглицеридов в лубе отмечается в период покоя (Рубчевская, Левин, 1982). При фитоценотическом стрессе было обнаружено значительное уменьшение количества жиров осенью в лубяной ткани ствола и корней, а также и в хвое деревьев. Таким образом, можно сделать вывод о том, что трофическое обеспечение ростовых процессов угнетенных деревьев значительно снижено, как за счет уменьшения содержания белков и крахмала, так и за счет основного запасного вещества хвойных – жиров.
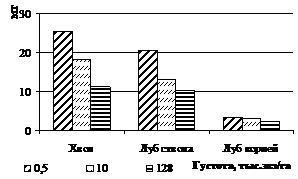
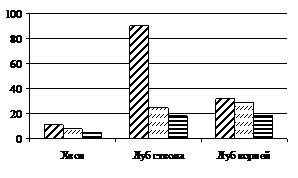 Рисунок 5 – Содержание триацилглицеридов (а) и водорастворимых фенольных соединений (б) в хвое (100 шт) и в одном см2 луба, приходящегося на один см3 камбиальной поверхности ствола и корней (мг) лиственницы сибирской в фитоценозах различной густоты
Рисунок 5 – Содержание триацилглицеридов (а) и водорастворимых фенольных соединений (б) в хвое (100 шт) и в одном см2 луба, приходящегося на один см3 камбиальной поверхности ствола и корней (мг) лиственницы сибирской в фитоценозах различной густоты
Накопление низкомолекулярных фенольных соединений в тканях хвойных является индикационным признаком низкотемпературного и водного стрессов (Судачкова и др., 1997). При фитоценотическом стрессе, однако, было обнаружено достоверное снижение содержания суммарного количества фенольных соединений в лубяной ткани деревьев лиственницы сибирской осенью (рис. 5б), а также и летом. Незначимые изменения содержания фенольных соединений при увеличении густоты посадки были отмечены в хвое осенью. Наибольшее снижение фенолов наблюдается в лубе ствола.
Согласно теории конкурентных отношений между синтезом белка и флавоноидов (Судачкова, 1977), при снижении уровня азотсодержащих соединений в отдельных структурах угнетенных деревьев можно было бы ожидать увеличения концентрации фенольных соединений. При распаде белка, что является неспецифической реакцией растения на любой стресс, наблюдается усиление образования фенолов, что является механизмом, сформировавшимся в процессе эволюции для блокирования токсических фенольных продуктов деградации белков. Возможно, при фитоценотическом стрессе такой механизм не осуществляется по причине низкой каталитической активности ферментов или малого количества продуктов распада, что подлежит дальнейшему изучению. Более вероятно, что снижение содержания фенольных соединений связано с уменьшением запасных веществ при фитоценотическом стрессе, поскольку отдельные соединения этого класса веществ выполняют резервную функцию. Так, гликозиды флавоноидов могут расщепляться, образуя агликон фенольной природы и гликозидный остаток, или полностью подвергаться катаболизму, вплоть до углекислоты в процессе дыхания (Запрометов, 1993). Кроме того, гликозидный остаток в гликозидах фенолов может служить своеобразной формой запасания сахаров (Chalker-Scott, 1999).
Снижение содержания фенольных соединений в тканях хвойных при фитоценотическом стрессе можно считать специфической реакцией, поскольку для изученных видов стресса, влияющих на древесные растения (низкотемпературный, осмотический, техногенный), характерно увеличение фенолов.
Выводы
Комплексный морфолого-анатомический анализ одновозрастных древостоев лиственницы сибирской показал, что фитоценотический стресс вызывает уменьшение размеров и массы структурных частей деревьев, ширины годичного кольца и приводит к изменению соотношения поздней и ранней древесины. Обнаружено, что толщина клеточных стенок поздней древесины снижается более интенсивно, чем ранней. Показано, что в загущенных посадках лиственницы сибирской ширина годичного кольца деревьев постепенно снижается, с возрастом почти не реагируя на изменение погодных условий.
Обнаружено снижение содержания общего и белкового азота,а также свободных аминокислот в отдельных органах деревьев.
Установлено, что запасы крахмала достоверно уменьшаются во всех органах и тканях деревьев лиственницы при фитоценотическом стрессе. При этом содержание моно- и олигосахаров изменяется в тканях деревьев неоднозначно. Характерно достоверное увеличение концентрации олигосахаров в лубяной ткани ствола. По-видимому, это связано с тем, что у деревьев нарушен процесс оттока и включения моно- и олигосахаров в полимерные соединения. Суммарное количество неструктурных углеводов, как правило, снижается в тканях деревьев из загущенных посадок.
Установлено, что в тканях деревьев лиственницы сибирской при фитоценотическом стрессе уменьшается содержание триацилглицеридов, которые являются основным энергетическим запасом хвойных. Трофическое обеспечение ростовых процессов угнетенных деревьев также значительно снижено за счет уменьшения содержания белков и крахмала.
Отмечено, что снижение содержания фенольных соединений в тканях лиственницы при увеличении густоты посадки может выступать в качестве биохимического индикатора фитоценотического стресса. Снижение содержания фенолов, по-видимому, связано с резервной функцией отдельных соединений этого класса веществ.
Библиографический список
Бузыкин, А.И. Густота и продуктивность древесных ценозов / А.И. Бузыкин, Л.С. Пшеничникова, В.Г. Суховольский. - Новосибирск: Наука, 2002. - 153 с.
Бузыкин, А.И. Рост и продуктивность молодняков разной густоты лиственницы сибирской / А.И. Бузыкин, Л.С. Пшеничникова // Лесное хозяйство, 2001. - № 5. - С. 34-35.
Вознесенский, В.Л. Определение сахаров по обесцвечиванию жидкости Фелинга / В.Л. Вознесенский [и др.] // Физиол. растений. - 1962. - Т.9, Вып.2. - С. 255-266.
Георгиевский, Н.П. Рубки ухода за лесом / Н.П. Георгиевский. - М.-Л.: Гослесбумиздат, 1957. - 142 с.
Давыдов, А.В. Влияние рубок ухода за лесом на механические свойства древесины / А.В. Давыдов, И.А. Беляев // Сб. ЦНИИЛС “Рубки ухода за лесом”. - Л.: Гослестехиздат, 1940. - С. 49-64.
Ермаков, А. И. Методы биохимического исследования растений / А.И. Ермаков [и др.]. - Л.: Колос, 1972. - 456 c.
Жуков, А.Б. Общелесоводственные мероприятия по повышению продуктивности лесов СССР. Проблемы повышения продуктивности лесов / А.Б. Жуков, А.Т. Савельев. - М.-Л.: Гослесбумиздат, 1958. - Т. 1. - 148 с.
Запрометов, М.Н. Фенольные соединения. Распространение, метаболизм и функции в растениях / М.Н. Запрометов. - Москва: Наука. - 1993. - 271 с.
Злобин, Ю.А. Оценка качества подроста древесных растений / Ю.А. Злобин // Лесоведение. - 1970. - №3. - С. 96-102.
Карпов, В.Г. Экспериментальная фитоценология темнохвойной тайги / В.Г. Карпов. - Л.: Наука, 1969. - 335 с.
Морозов, Г.Ф. Учение о лесе. Вып. 1. Введение в биологию леса. / Г.Ф. Морозов. - С.-Петербург, 1912. - 83 с.
Плешков, Б.П. Практикум по биохимии растений / Б.П. Плешков. - М.: Колос, 1968. - 183 с.
Рубчевская, Л.П. Об ацилглицеролах камбиальной зоны лиственницы сибирской / Л.П. Рубчевская, Э.Д. Левин // Химия древесины. - 1982. - № 1. - С. 107-109.
Смолистые вещества древесины и целлюлозы, их роль в целлюлозно-бумажной промышленности / Под ред. И.А. Нагродского. - М.: Лесная промышленность, 1968. - 349 с.
Судачкова Н.Е. Метаболизм хвойных и формирование древесины / Н.Е. Судачкова. - Новосибирск: Наука. Сибирское отделение, 1977. - 231 с.
Судачкова, Н.Е. Биохимическая адаптация подроста хвойных к условиям существования под пологом леса / Н.Е. Судачкова, Л.И. Балмаева // Лесоведение. - 1974. - №4. - С. 16-23.
Судачкова, Н.Е. Биохимические индикаторы стрессового состояния растений / Н.Е. Судачкова [и др.]. - Новосибирск: Наука. Сиб. предприятие РАН, 1997. - 176 с.
Судачкова, Н.Е. Физиология подроста кедра / Н.Е. Судачкова, Е.Я. Расторгуева, Р.А. Коловский. - М.: Наука, 1967. - 122 с.
Судницына, Т.Н. Азотное питание сосны в культурах различной густоты посадки / Т.Н. Судницына // Лесоведение. - 1967. - №2. - С. 67-73.
Сукачев, В.Н. О внутривидовых и межвидовых взаимоотношениях среди растений / В.Н. Сукачев // Сообщения Института леса. - М.: Изд-во АН СССР. - 1953. - С. 5-44.
Татаринов, В.В. Экспериментальный анализ факторов, регулирующих численность и рост подроста сосны в сложных сосняках Беловежской пущи / В.В. Татаринов // Лесоведение. - 1972. - №4. - С. 3-12.
Тимофеев, В.П. Осветления и прочистки / В.П. Тимофеев. - М.: Гослесбумиздат, 1950. – 87с.
Chalker-Scott, L. Environmental Significance of Anthocyanins in Plant Stress Responses / L. Chalker-Scott // Photochem. Photobiol. - 1999. - V. 70. - P. 1–9.
Humphreys, F.R. A method for determination of starch in wood / F.R. Humphreys, J. Kelly // Anal. Chem. Acta. - 1961. - V. 24. N 1. - P. 66-70.
Methods of enzymatic food analysis / by H.U. Bergmeyer (Editor). - Mannheim GmbH., 1984. - P. 75-77.
Поступила в редакцию 10 марта 2008 г.
Принята к печати 27 августа 2008 г. |


