|
|
|
Rus/Eng |
|
||||||||||||
|
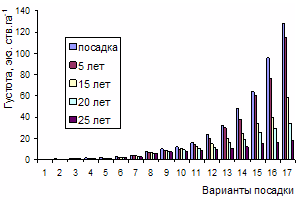
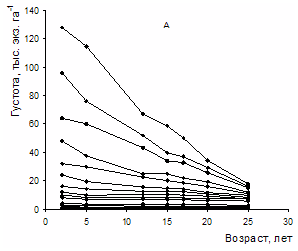
"Хвойные бореальной зоны" 2008г.,№3-4, с. 244-249 Естественное изреживание разногустотных молодняков в экспериментальных посадках сосны Бузыкин А.И., Пшеничникова Л.С., Собачкин Д.С., Собачкин Р.С. Институт леса им. В.Н.Сукачева СО РАН В многовариантных по густоте посадках сосны показано естественное изреживание ценозов за 25 лет. Оригинальные натурные данные позволили корректно оценить роль густоты в регулировании собственной плотности древесных ценозов. Рост древесных ценозов сопровождается уменьшением их густоты с возрастом в процессе дискриминационного и стохастического отпадов. Если по всему густотному ряду крайние значения густоты начальной посадки различались в 256 раз, то в 25-летних ценозах – в 78 раз. Наиболее интенсивный дискриминационный отпад в течение 25 лет происходит в ценозах с вариантами густоты в пределах 24-128 тыс. экз. га-1 . Связь отпада деревьев по группам густот и густотному ряду с возрастом, несмотря на большую вариабельность, является очень тесной. Изменение густоты и роста в процессе изреживания с возрастом для ценозов разной плотности имеют свои особенности и зависят от начальной густоты (густоты посадки) ценоза: в свою очередь ценоз сам выступает регулятором собственной плотности. Ключевые слова: густота насаждений, отпад деревьев, естественное изреживание ценозов This paper deals with self-thinning of forest plantation over a 25-year period depending on planting density. Our experimental field data enabled correct assessment of the importance of stand density for stand self-thinning. Stand density was found to decrease with increasing stand age due to competition-caused and stochastic tree mortality. While the initial lowest and highest plantation densities differed 256 times, the density difference dropped to 78 times in 25-year-old stands. Forest stands with densities ranging 24 to 128 thousand trees per hectare showed the highest competition-caused tree die-back. Stand age appeared to have a strong relationship with tree mortality at any stand density. Forest stand growth and self-thinning exhibited stand density-specific characteristics. On the other hand, a forest stand was determined to regulate its own density. Key words: stand density, stand density, self-thinning of forest plantation Введение Естественные древесные ценозы в однородных лесорастительных условиях и в пределах одного возраста очень вариабельны по густоте. Фактор густоты в лесоэкологических, лесоводственных и таксационных исследованиях существенно усложняет получение однозначной информации о структуре и продуктивности лесных экосистем вследствие того, что адекватная оценка роли густоты возможна на основе анализа многовариантных густотных рядов. Использование в исследованиях несистемных, а также случайных или отдельных вариантов густоты обычно не приносят значимого результата сверх того, что густоту древесных ценозов надо как-то учитывать (Левин, 1966; Рубцов, 1969; Мартынов, 1974; Рубцов и др., 1976; Бузыкин и др., 2002). Можно ожидать, что лишь многовариантные по густоте ряды, характеризующие густоту в широких пределах – от роста деревьев свободного стояния до загущенных и очень густых древесных ценозов, имеющих десятки и сотни тыс. особей на га – являются наиболее информативными и адекватными объектами исследования роли плотности древесных ценозов (Бузыкин, Пшеничникова, 1999). ОБЪЕКТЫ И МЕТОДИКА ИССЛЕДОВАНИЙ В 1982 году сотрудниками Института леса СО РАН под руководством А.И. Бузыкина в подзоне южной тайги (Большемуртинский лесхоз Красноярского края) на серых лесных почвах в однородных лесорастительных условиях плакора созданы посадки сосны обыкновенной, лиственницы сибирской, ели сибирской из 18 вариантов густоты: 0,5, 0,75, 1, 1,5, 2, 3, 4, 6, 8, 10, 12, 16, 24, 32, 48, 64, 96, 128 тыс. экз. га-1, т.е. крайние варианты густоты посадки различались в 256 раз. Для посадки по квадратной схеме использовались двухлетние сеянцы. На участке каждого варианта густоты высаживалось не менее 500 растений; причем для вариантов с густотой от 48 до 128 тыс. экз. га-1 число посадочных мест на участках увеличивалось соответственно на 10-40%. Площадь каждого участка определялась густотой варианта, по конфигурации она близка к квадратной. Варианты примыкали друг к другу без разрывов, образуя целостные блоки посадок каждой породы, отделяемые друг от друга пятиметровым разрывом. Общая площадь экспериментальных посадок составила 17 га. РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ Особый интерес представляет изменение густоты в процессе естественного изреживания. Рассмотрим динамику густоты сосновых посадок и образуемых ими ценозов за 25 лет. На рис. 1 представлена начальная густота посадки и ее состояние в 5, 15, 20 и 25-летнем возрасте.
Рисунок 1 - Начальная и последующая густота в 5, 15, 20 и 25-летнем возрасте по вариантам плотности В первые 3 года определялись приживаемость высаженных саженцев и их послепосадочная сохранность.
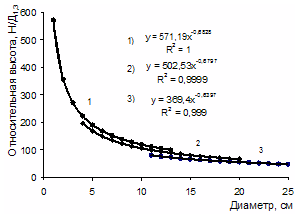
Рисунок 2 - Относительная высота деревьев в 25-летних ценозах разной густоты посадки: 1 – 128, 2 – 4, 3 – 0,5 тыс. экз.-1
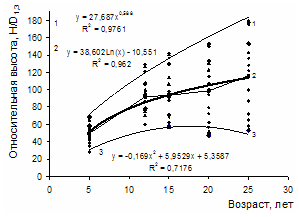
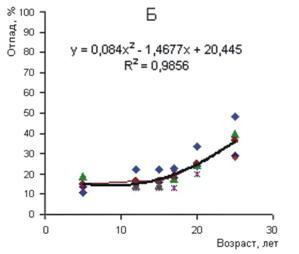
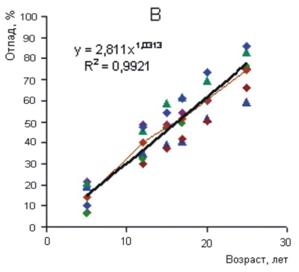
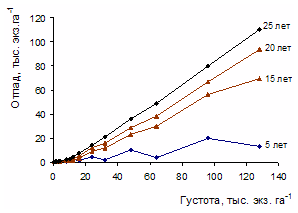
Рисунок 3 - Изменение средней относительной высоты разногустотных ценозов с возрастом: 1-3 – верхний и нижний пределы амплитуды поля рассеивания средних значений относительных высот разногустотных ценозов; 2 – средняя относительная высота для всего густотного ряда Изменение густоты в процессе изреживания с возрастом для ценозов разной плотности имеет свои особенности. На рис. 4 представлен относительный отпад в 3-х совокупностях ценозов низкой, средней и высокой густоты. За период наблюдений вариабельность отпада с возрастом в пределах густотного ряда характеризуется в диапазоне от значительной до очень большой (Дворецкий, 1971). Наибольшая относительная величина суммарного отпада в 5-летнем возрасте оказалась в ценозах низкой густоты – 20-35%; в группах ценозов с начальной густотой 4-16 и 24-128 тыс. экз. га-1 отпад находился в пределах 10-20 и 5-20%. В 25-летнем возрасте по группам густот сохранилось соответственно 40-65, 45-70 и 15-40% деревьев от числа высаженных. В целом по группам густот и всему густотному ряду амплитуда суммарного отпада с возрастом возрастает, а среднего и текущего отпада уменьшается (рис. 5).
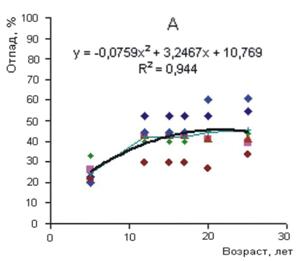
Рисунок 4 - Суммарный отпад в разногустотных ценозах разного возраста. Густота: А – 0,5-3, Б – 4-16, В – 24-128 тыс. экз. га-1 Густота представляет собой важный структурно-функциональный показатель ценоза, с ней связаны жизненно важные ресурсы среды, приходящиеся на одну особь. Если бы естественные механизмы регуляции однозначно приводили численность ценоза в соответствие с экологическими условиями и ресурсами среды, то задача выявления целевой и оптимальной густоты или ее экологической роли решалась бы сравнительно просто. Однако функционирование механизмов регуляции численности осложняется приспособительными реакциями древесных растений к экологической и фитоценотической среде. Особенно широкая амплитуда морфофизиологической адаптации свойственна хвойным. В пределах ценопопуляции растения одного и того же возраста, как уже отмечалось, по своей фитомассе могут различаться в 102—103 и более раз.
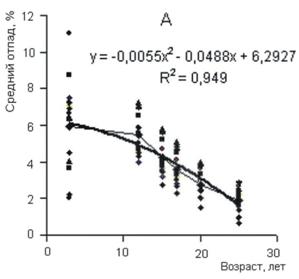
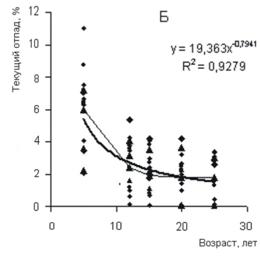
Рисунок 5 - Изменение ежегодных среднего (А) и текущего (Б) отпада разногустотных ценозов с возрастом В древесных ценозах в процессе их роста и развития функционируют два процесса или механизма регуляции густоты ценоза и морфоструктуры древесных растений. Один из них регулирует численность деревьев и стремится привести ее в соответствие с ресурсами среды и емкостью биологического круговорота. Поскольку в основе этого механизма лежат экологические факторы, то он, в сущности, экологически детерминирован. Другой процесс направлен на стабилизацию численности деревьев и поддержание ее на высоком уровне за счет приспособительных реакций особей к складывающейся фитоценотической ситуации. Основу механизма ответных реакций растений на давление внешней и фитоценотической среды составляют физиолого-биохимические процессы, т. е. они детерминированы биологически (эволюционно и генетически). Так как эти механизмы связаны с процессом роста ценоза и контролируются средой, то их разделение в известной степени условно. По существу они представляют разные стороны одного процесса регуляции густоты древесных ценозов:
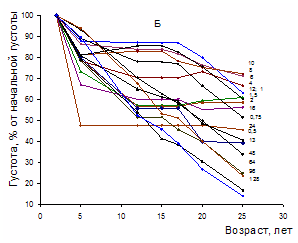
Рисунок 6 - Изменение абсолютной (А) и относительной (Б) густоты сосновых ценозов с возрастом. Цифрами обозначена густота посадки в тыс. экз. га-1 Фактически элиминация наступает тогда, когда ее уже не в состоянии сдержать морфофизиологическая перестройка растения, т.е. элиминирующее давление превышает норму реакции растений на это давление. При этом регуляция численности деревьев существенно запаздывает по сравнению с ростовой реакцией на недостаток тех или иных ресурсов и воздействие фитоценотических и экологических факторов.
Рисунок 7 - Отпад деревьев разногустотных ценозов в разном возрасте Численность и размеры деревьев в реальном ценозе нужно рассматривать как результат существования в нем подвижного единства экологически и биологически детерминированных механизмов регуляции плотности ценопопуляции и реакции древесных растений через ростовые процессы и соответствующие им размеры особей на экологическую и фитоценотическую обстановку. В условиях высокой плотности в процессе роста ценоза первый механизм направлен на элиминацию лишних особей, второй — на выживание возможно большего числа индивидуумов и поддержание высокой численности за счет депрессии роста, снижения размеров особей, перестройки их морфоструктуры и физиолого-биохимических процессов. В последнее время дендрофизиологами и биохимиками получены весьма интересные данные зависимости метаболизма древесных растений от густоты, обусловленные влиянием ценотического стресса на рост деревьев при конкурентных взаимодействиях в условиях естественного загущения и загущенных посадок (Плаксина и др., 2001; Судачкова, 1998 и др.). Выделение результатов действия этих механизмов в "чистом" виде представляет значительные трудности. Есть основания полагать, что биологически детерминированная реакция древостоя на загущение без анализа густотных рядов обычно принимается за экологически обусловленные густоту и рост древостоев. ЗАКЛЮЧЕНИЕ Фактическая плотность древесных ценозов и ее густотные градиенты в публикациях по лесной экологии и лесоведению почти не рассматриваются, хотя они определяют размер и объем ресурсов среды, включая пространство роста, используемые каждой особью в процессе своего роста и развития. В лесоэкологических исследованиях целесообразно оперировать прямыми показателями плотности некоторого множества разногустотных объектов, представляющих густотный ряд, для того чтобы знать степень зависимости полученных результатов от плотности древесных ценозов – атрибута лесных экосистем. БИБЛИОГРАФИЧЕСКИЙ СПИСОК Бузыкин, А.И. Влияние густоты на морфоструктуру и продуктивность культур сосны / А.И. Бузыкин,Л.С. Пшеничникова // Лесоведение. – 1999. - № 1. - С. 38-43. Поступила в редакцию 25 апреля 2008 г.
|
|||||||||||