|
"Хвойные бореальной зоны" 2008г.,№1-2, с. 16-20
Структура и функции грибного комплекса лесного биогеоценоза
Стороженко В.Г.
Институт лесоведения РАН
143030 с. Успенское, Одинцовский р-н, Московская обл.; e-mail: root@ilan.msk.ru
На основе многолетних исследования структур фитоценозов и грибных дереворазрушающих комплексов лесов различного происхождения на Русской равнине оценена роль и функции грибов в генезисе лесных сообществ. Рассмотрены трофотопические, консортивные и функциональные взаимоотношения грибной биоты и автотрофов в лесном биогеоценозе. Определены функциональные задачи грибной биоты дереворазрушающего комплекса лесных сообществ.
Ключевые слова: Русская равнина, дереворазрушающие грибы, фитоценоз
The author made a survey of the structures of phytocenosis and fungi wood destroying complexes in the different origin forests on the Russian plain. The author also estimated the role and functions of wood destroying complexes fungi in the wood communities genesis. There were also examined tropho-topic, consortive and functional interaction of fungi biota and autotrophs in the forest of different dynamic characteristics. Functional objectives of the wood destroying complexes fungi biota in the woods formation have been also defined by the author.
Key words: Russian plain, fungi wood destroying complexes, phytocenosis
Введение
Для лесных насаждений искусственного происхождения и лесного хозяйства многих стран мира, в том числе России, существенное экосистемное и хозяйственное значение имеет постоянно фиксируемое очаговое распространение некоторых дереворазрушающих грибов, среди которых наиболее вредоносными можно признать корневую губку (Heterobasidion annosum (Fr.) Bref.), некоторые виды рода Armillaria (A. mellea, A.borealis, A. ostoyae) – опенок или некоторые другие возбудители, вызывающие очаговое поражение лесов. Половину прошлого века ведущие научно-исследовательские центры России, республик Прибалтики, Украины, Казахстана и многих зарубежных стран мира изучали возможности борьбы с этими грибами. На поиски этой борьбы тратились и продолжают тратиться огромные деньги, людские и временные ресурсы. В итоге этих широчайших по масштабам работ мировая научная общественность вынуждена была заключить, что никакие средства борьбы не могут остановить распространение этих возбудителей и предотвратить развал ими искусственно создаваемых древостоев. В то же время почти всегда приводились сведения о том, что в лесах естественного происхождения, тем более коренных разновозрастных очагов этих возбудителей, нет, хотя сами грибы присутствуют в составе микоценозов лесных сообществ.
По нашему убеждению, такая ситуация сложилась из-за того, что в лесной биогеоценологии, микологии, в фитопатологии и лесном деле не утвердилось представление о структурной иерархии грибов в лесном сообществе, о функциональных задачах групп и комплексов грибов, выработанных в их эволюционной коадаптационной истории с высшими растениями.
А между тем анализ структурного и функционального устройства лесного микоценоза позволяет объяснить и понять причины такого поведения грибов и при желании определить пути воздействия на них. В настоящей статье делается попытка на основе рассмотренных структурных и функциональных параметров грибных сообществ лесных биогеоценозов и изученных закономерностей поведения грибов в лесах различных лесоводственных характеристик (Стороженко и др., 1992, Стороженко, 2002, 2005, 2005а, 2007) оценить роль грибной биоты, а в нашем случае комплекса дереворазрушающих грибов, в генезисе лесных биогеоценозов.
ПРОГРАММА И МЕТОДЫ ИССЛЕДОВАНИЙ
Эволюционное устройство структур и функций грибной биоты лесов может быть понято и объяснено только при анализе не затронутых антропогенным воздействием девственных лесных сообществ различных коренных формаций в сравнительном анализе с лесами условно-коренными, искусственного происхождения и подверженными различного рода антропогенным воздействиям. Это положение лежало в основе подбора площадей лесов и методик их анализа при наших исследованиях. Изучались структуры фитоценозов – возрастные, горизонтальные, валежные, возобновительные; грибные дереворазрушающие комплексы биотрофов на живых деревьях и ксилотрофов, разлагающих древесный отпад. Определялся видовой состав дереворазрушающих грибов, их распространенность в древостоях, пораженность древостоев, соотношение видов грибов в структурах микоценозов.
Всего было заложено более 500 постоянных и временных пробных площадей.
Исследования проводились в лесах всех природных зон и основных коренных формаций на Русской равнине. Для сравнительной оценки поведения грибов изучались структуры грибных сообществ условно-коренных и искусственных лесов различных лесоводственных параметров.
Результаты исследований
Прежде чем рассматривать структурные и функциональные характеристики грибного сообщества лесного биогеоценоза, необходимо прийти к пониманию того, что грибное сообщество действительно является ценотической структурой этого лесного сообщества, а не случайным сбором грибов в нём. В наших более ранних публикациях эти доказательства приведены и подробно описаны (Стороженко и др., 1992; Стороженко, 1994, 2000, 2002, 2005, 2007 и др.). В общих чертах повторим, что в статусе ценотической структуры в составе, в частности, лесного биогеоценоза грибная биота может рассматриваться по присущим ей характеристикам макротаксономического строения филемы органического мира (Кусакин, Дроздов, 1994, 1998), где грибы трактуются как царство (Fungi), наравне с царствами растений (Plantae) и животных (Animalia) (Гарибова, 1999; Гарибова, Лекомцева, 2005), по присутствию в структуре грибной биоты морфологического, экологического и функционального строения (Стороженко и др., 1992) и по роли грибной биоты в осуществлении процессов круговорота веществ и энергии в природных циклах. Все эти качества позволяют безальтернативно причислять грибное сообщество к ценотическим структурам лесного биогеоценоза, по значению равноценного другим ценотическим структурам, в том числе и фитоценозу (Стороженко, 1992, 2007 и др.).
В структурах лесного фитоценоза грибная биота имеет вполне определенное, эволюционно закономерно обусловленное разделение видового состава грибов, принадлежащих к различным группам по таксономическому положению и способу питания на различных частях растений. Это разделение, безусловно, связано с морфологической структурой растений и ярусов лесного полога (рис.1).
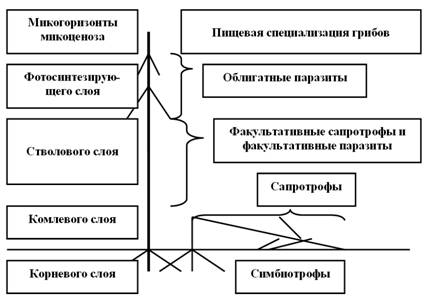
Рисунок 1 – Схематическое изображение структуры лесного микоценоза
Представленная на рисунке схема отражает закономерности коэволюционного сосущество-вания биоты грибов и растений в лесном биогеоценозе. В этой схеме не может быть перестановок или замены комплексов грибов. Такая структура имеет весьма жесткий трофотопический каркас, в котором каждая группа грибов занимает свою экологическую нишу, определяемую морфоструктурой, состоянием и положением субстрата. Именно так эволюционно построена структура лесного микоценоза, при которой он занимает все пространство леса и в котором все грибы взаимообусловленно существуют без какого-то вреда для него в соотношениях, необходимых и достаточных для выполнения эволюционно определенных функций.
Структурное морфологическое и экологическое устройство грибной биоты насыщено в такой же степени закономерно распределенными консортивными связями взаимного функционирования комплексов грибов в микоценозе и в фитоценозе (рис. 2).
Группы грибов облигатных паразитов осуществляют свой жизненный цикл в основном на фотосинтезирующих, проводящих, неодревесневших активных тканях растущих растений, убивая их, и эта биомасса древесного опада затем перерабатывается сапротрофами – консорция консументов. К консорции консументов относятся также симбиотрофы.
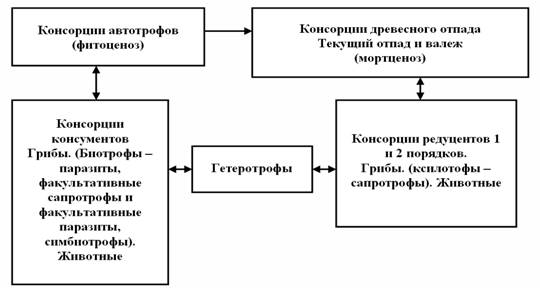
Рисунок 2 – Схематическое изображение консорций лесного биогеоценоза
Грибы, обладающие факультативными свойствами с разной степенью паразитической и сапрофитической активности, также поселяются на живых деревьях, поражают в большинстве своём менее активные ткани растений, вызывая гнили разных типов стволов, ветвей, корней и доводят деревья до гибели. Этот процесс гораздо медленнее и избирательнее, чем при поражении растений облигатными паразитами - также консументы. Многие из грибов этой группы способны долгое время функционировать на отмерших деревьях или их фрагментах, переходя в состав редуцентов.
Комплекс сапротрофов изначально предназначен для переработки отмерших растений фитоценоза или их частей на живых деревьях, древесного отпада и опада, отмерших плодовых тел самих грибов и другой биомассы биогеоценозов. Эта группа грибов относится к редуцентам 1 и 2 порядков, некротрофам.
Все эти грибы в лесах эволюционного развития, тем более девственных, сбалансированы по числу и представленности видов и комплексов различной пищевой специализации в составе своих микоценозов. В таких лесах колебания состава грибной биоты зависят и соответствуют только сукцессионному положению лесного сообщества. Нас в первую очередь интересует группа грибов биотрофов дереворазрушающего комплекса, обладающих факультативными свойствами, и ксилотрофов сапротрофного комплекса. Обширными и долговременными исследованиями структур фитоценозов и грибных комплексов лесов на Русской равнине определено, что грибная биота имеет закономерно обусловленные параметры поражения древостоев фитоценозов как по количественным (уровни и объемы поражения), так и по качественным (соотношение видов грибов различной активности по отношению к автотрофам). Это значит, что каждому этапу или фазе формирования естественно развивающегося лесного сообщества определенного формационного состава в определенных экологических и зонально-климатических условиях, при определенных структурных и лесоводственных характеристиках соответствует определенный состав и структура грибов. В наибольшей степени это относится к грибам факультативной группы. Важность этой группы грибов состоит в том, что именно эти грибы осуществляют поражение деревьев, вызывают гнили стволов, ветвей и корней, приводят к ослаблению их механических свойств, буреломам, ветровалам или постепенному усыханию на корню. Постоянно в лесном сообществе происходит выборка, изьятие определенного количества деревьев из состава древостоя и перевод их в категорию древесного отпада. Таким образом, комплекс дереворазрушающих биотрофных грибов активно участвует в формировании структур фитоценозов и поддержании баланса в процессах накопления и отпада биомассы в лесных сообществах. Определено и доказано, что существует на уровне закономерности вполне определенный баланс между стадией сукцессионного развития лесного сообщества и структурной организацией биоты грибов и, в частности, грибов дереворазрушающего биотрофного комплекса (Стороженко, 2007) (рис.3).
Наивысшие показатели этого баланса достигаются в лесах климаксовых динамических характеристик. В лесах любых других динамических показателей этот баланс сдвигается в область преобладания видов грибов, с большими или меньшими факультативными свойствами паразитизма или сапрофитизма. В состав консументов этой группы грибов входят и виды, обладающие способностью к очаговому распространению, такие как корневая губка (Heterobasidion annosum (Fr.) Bref.) и виды рода Armillaria (A. mellea, A.borealis, A. ostoyae). Однако никогда в коренных разновозрастных, тем более девственных, сбалансированных по структурным и функциональным характеристикам лесах не бывает очагов этих возбудителей. Их активность контролируется общим антагонистическим полем сбалансированного микоценоза. .
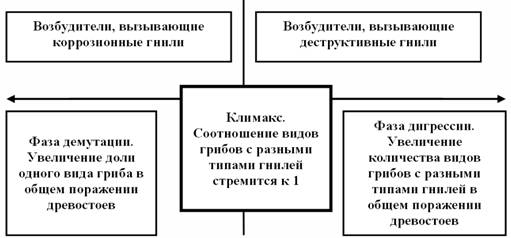
Рисунок 3 – Схематическое изображение функциональных закономерностей участия дереворазрушающих грибов, вызывающих разные типы гнилей, в общем поражении коренных лесов
Дереворазрушающие грибы биотрофного комплекса выполняют в определённой степени функцию эндогенного механизма регуляции формирования структур фитоценоза. Именно поэтому, если возникает дисбаланс в структурах фитоценоза немедленно вступает в действие этот регуляционный механизм. В соответствии с изменением структур фитоценоза изменяется состав биотрофных грибов, уровни поражения ими древостоев, величина древесного отпада. В древостоях искусственного происхождения, создаваемых, как правило, без учета основных закономерностей формирования устойчивых лесов (Стороженко, 2002, 2005, 2007), факультативные виды дереворазрушителей, обладающих способностью к быстрому наращиванию патогенности, такие как обозначенные выше виды, реализуют эту способность, принимая непосредственное участие в формировании более устойчивых лесных сообществ. Это участие проявляется в стремлении как можно быстрее разрушить неустойчивые структуры сообщества для их последующей перестройки через смену пород в более устойчивые. Примеры такого поведения грибов зафиксированы ещё в середине Х1Х века в Южной Баварии. В ХХ веке очаговое поражение лесных культур стало приобретать масштабы эпифитотий во многих странах мира, в том числе в России (Heterobasidion ..., 1998; Павлов, 2007). Пик активности борьбы с очаговым поражением искусственных лесов хвойных формаций пришёлся на середину ХХ века, когда во многих странах мира создавались и испытывались различные механические, лесохозяйственные, химические и биологические методы профилактики, локализации очагов и борьбы с распространением перечисленных выше грибов. Однако к концу прошлого века многие исследователи, работавшие в этом направлении, вынуждены были прийти к заключению, что опыт многих десятилетий активной борьбы с корневой губкой и опенком не дал сколько-нибудь ощутимых результатов. Эволюционный механизм регуляции оказывался каждый раз сильнее искусственных мер борьбы. Приходится констатировать, что единственный путь избежать распространения этих и других возбудителей очагового поражения – максимально приблизить структуры лесов к модели коренной формации по лесоводственным характеристикам.
Таким образом, если суммировать все изложеннные выше закономерности формирования комплексов дереворазрушающих грибов как части микоценоза лесного сообщества, можно оценить функциональные задачи грибной биоты, в нашем случае комплексов дереворазрушающих грибов. Их важнейшей функциональной задачей является регуляция структур фитоценозов в процессах сукцессионного движения лесного сообщества к состоянию наибольшей сбалансированности всех его ценотических структур. Решение этой эволюционно определенной функциональной задачи достигается через реализацию ряда побочных функций – деструктивной функции, осуществляемой грибами факультативной и сапротрофной групп при поражении живых деревьев и древесного отпада, возможности изменять в определенных пределах трофическую специализацию, адаптационную способность при изменениии свойств субстрата, которые проявляются на уровне отдельных деревьев и в целом лесного сообщества.
Именно эту основную функцию, на наш взгляд, следует рассматривать как один из важнейших гетеротрофных эволюционных механизмов, участвующих в эволюционном движении сообществ по пути их сукцессионного развития. Этот механизм объединяет процессы ослабления деревьев, поражения древостоев в количественных и качественных показателях, накопление древесного отпада и скорость его разложения сапротрофным комплексом в единый сбалансированный процесс, отражающий структурные и динамические характеристики лесного сообщества.
В практическом применении такое понимание роли и функций грибной биоты в формировании лесов позволяет избежать огромных затрат на поиск и осуществление средств борьбы с очаговым поражением лесов и создавать леса, устойчивые к различным неблагоприятным факторам воздействия.
Основные критерии создания и формирования устойчивых лесов рассмотрены в некоторых публикациях ранее (Стороженко и др., 1992, 2005, 2007; Павлов, 2007).
Заключение
1. Грибное сообщество лесного биогеоценоза является его ценотической структурой, что определяется присущими ему характеристиками макротаксономического строения филемы органического мира по присутствию в структуре грибной биоты морфологического, экологического и функционального строения и по роли грибной биоты в осуществлении процессов круговорота веществ и энергии в природных циклах.
2. В структурах лесного фитоценоза грибная биота имеет вполне определенное, эволюционно закономерно обусловленное разделение по видовому составу грибов, принадлежащих к различным группам по таксономическому положению и способу питания на различных частях растений.
3. Структурное морфологическое и экологическое устройство грибной биоты насыщено в такой же степени закономерно распределенными консортивными связями взаимного функционирования комплексов грибов в микоценозе и фитоценозе.
4. В целом сбалансированный по видовому составу и пищевой специализации микоценоз устойчивых коренных лесов, в том числе комплексы дереворазрушающих грибов биотрофной и ксилотрофной групп, соответствует динамическим характеристикам биогеоценоза, имеет вполне определенную структуру и иерархическую подчиненность видов, осуществляя контроль за текущим отпадом нужного количества деревьев из состава древостоев.
5. Важнейшей функциональной задачей биотрофного (в том числе дереворазрушающего) комплекса грибов является регуляция структур фитоценозов в процессах сукцессионного движения лесного сообщества к состоянию наибольшей сбалансированности всех его ценотических структур.
Проведенные исследования имеют существенное прикладное значение, так как позволяют правильно расставить акценты в понимании роли и функций грибной биоты лесов, прогнозировать их будущее развитие, возможность и направление лесохозяйственных мер создания устойчивых лесных сообществ.
Библиографический список
- Гарибова, Л.В. Обзор и анализ современных систем грибов / Л.В. Гарибова.- Петрозаводск, 1999. – 28 с.
- Гарибова, Л.В. Основы микологии / Л.В. Гарибова, С.Н. Лекомцева. – М.; 2005.- 220 с.
- Кусакин, О.Г. Филема органического мира / О.Г. Кусакин, А.Л. Дроздов. – СПб., 1994.- Ч.-1. 281 с.
- Кусакин, О.Г. Филема органического мира / О.Г. Кусакин, А.Л. Дроздов. –. СПб., 1998.- Ч.-2. 358 с.
- Научные основы устойчивости лесов к дереворазрушающим грибам / В.Г. Стороженко, [и др.]. – М.: Наука. 1992. 221 с.
- Павлов, И.Н. Техногенные и биотические механизмы деструкции коренных лесов юга Сибири и их восстановление: автореф. дис…. д-ра биол. наук / И.Н. Павлов.- М., 2007.- 43с.
- Стороженко, В.Г. Гнилевые фауты коренных лесов Русской равнины / В.Г. Стороженко. – М.: Изд-во ВНИИЛМ, 2002.- 156 с.
- Стороженко, В.Г. Грибные дереворазрушающие комплексы в генезисе еловых биогеоценозов : автореф. дис…. д-ра биол. наук / В.Г. Стороженко. – М.- 1994.- 42 с.
- Стороженко, В.Г. Понятия «больного» и «здорового» лесного сообщества / В.Г. Стороженко // Материалы 6-ой международной конференции «Проблемы лесной фитопат. и микол.» М.- Петрозаводск, 2005. -С. 311-316.
- Стороженко, В.Г. Разделение лесов по градациям устойчивости / В.Г. Стороженко // Материалы 6-ой международной конференции «Проблемы лесной фитопат. и микол.» М.- Петрозаводск, 2005. -С. 317-319.
- Стороженко, В.Г. Стратегии и функции грибных сообществ лесных экосистем / В.Г. Стороженко // Грибные сообщества лесных экосистем. – М.-Петрозаводск: Изд-во Карельского НЦ РАН, 2000.- С. 37-42.
- Стороженко, В.Г. Устойчивые лесные сообщества / В.Г. Стороженко. – М.- Изд-во «Гриф и К°»., 2007.- 190 с.
- Heterobasidion annosum. Biology, Ecology, Impact and Control.- New-York. CAB International, 1998.- 589 p.
|


