|
"Хвойные бореальной зоны" 2008г.,№1-2, с. 9-15
Сукцессионные процессы в лесных сообществах: модели фазовых переходов
Исаев А.С.*, Суховольский В.Г.**, Бузыкин А.И.**, Овчинникова Т.М **
*Центр по проблемам экологии и продуктивности лесов РАН
117997,Москва, ул. Профсоюзная, 84/32
**Институт леса им. В.Н.Сукачева СО РАН
660036 Красноярск, Академгородок, 50; е-mail: soukhovolsky@nm.ru
Работа посвящена разработке моделей, описывающих сукцесссионные переходы в лесных сообществах как фазовые переходы первого и второго рода. Рассматриваются изменения состава древесных ценозов под воздействием природных и антропогенных факторов. Предложена и верифицирована математическая модель последовательного перехода древесных ценозов из одного сукцессионного состояния в другое. Показано, что модель хорошо согласует с данными натурных наблюдений. Предлагаемый подход к моделированию сукцессионных процессов позволяет определить критические значения фитомассы насаждения, по достижению которой начинается сукцессионный переход.
Ключевые слова: лесные насаждения, фитомасса, сукцессии, модели, фазовые переходы
The work is devoted to development of model describing succession in the forest communities as first and second order phase transitions. Changes in the species composition of wood communities caused by natural and anthropogenic factors are considered. The math model of sequential succession transition in the wood communities is proposed and verified. It has been shown that the model is in good agreement with the actual data. The proposed approach allows establishing critical values of stand phytomass at which the succession transition begins.
Key words: forest stands, phytomass, successions, models, phase transitions
Работа выполнена при поддержке РФФИ (гранты №№ 05-04-49360 и 07-04-96804).
Введение
Процессы, происходящие в лесных экосистемах, весьма сложны и их теоретический анализ достаточно затруднен в связи с большим числом особей в экосистемах, значительным числом взаимодействий разного типа и сложной структурой размещения особей. В связи с этим при описании этих процессов обычно используются определенные упрощающие предположения. Одним из таких упрощающих предположений является понятие сукцессии. Лесные сукцессии в лесоведении и лесной биогеоценологии обычно характеризуют процесс смены древесных пород – эдификаторов и субэдификаторов (Сукачев, 1964). Сукцессионные процессы можно описывать как изменяющиеся во времени процессы, и тогда для описания таких процессов необходимы модели, учитывающие «кинетику» изменения состава насаждения, густоты и запаса фитомассы растений, либо как последовательность дискретных состояний ценоза. В этом случае необходимо определить набор отдельных состояний насаждения и последовательность смены одного состояния на другое (сукцессионный ряд). Обычно набор таких состояний (сукцессий) и переходов из одного состояния в другое (сукцессионных переходов) определяется экспертно, в связи с чем в литературе существует значительный разнобой в определении сукцессионной динамики конкретных ценозов (Александрова, 1964; Миркин, Розенберг
1978). Возможность реализации тех или иных сукцессионных переходов обычно определяют, исходя из имеющихся наблюдений сукцессионных процессов на данной территории. Однако несопоставимость характерных времен сукцессии (десятки и сотни лет) с характерными временами наблюдений за лесными территориями, сложность и субъективность процесса построения генетических рядов сукцессий затрудняет анализ сукцессионных процессов в лесных ценозах.
Для описания сукцессионных процессов в настоящей работе используются представления об экологических фазовых переходах (Soukhovolsky, 2007). Под фазой в физических системах понимается часть системы, однородная по своему составу и свойствам (Брус, Каули, 1984; Ландау, Лифшиц, 1964). При фазовом переходе физические свойства вещества изменяются (Ландау, Лифшиц, 1964; Дмитриев, 2004). В физических системах выделяются фазовые переходы первого и второго рода (Ландау, Лифшиц, 1964). Для фазового перехода первого рода в физических системах характерно наличие резких скачков плотности вещества, для фазового перехода второго рода свойства системы (такие как теплоемкость, намагниченность и др.) изменяются непрерывно.
В экологических системах развитие растительного сообщества также достаточно часто сопровождается резкими изменениями и величин фитомасс отдельных растений, и величиной общей фитомассы сообщества. В частности, такие резкие изменения (сукцессии) происходят в процессах смены сообщества травянистых растений на сообщество древесных растений. При переходе сообщества из состояния с преобладанием травянистых растений в состояние с преобладанием древесных растений достаточно быстро (за несколько лет) резко изменяется и суммарная фитомасса сообщества, и фитомассы растений травянистых и древесных видов. Фитомасса травянистых сообществ в среднем составляет несколько десятков тонн на гектар, тогда как фитомасса древесного насаждения – величина порядка сотен тонн на гектар (Замолодчиков и др., 2005).
При сукцессионных переходах другого типа происходит замена деревьев лиственных пород (березы, осины) на хвойные. При этом доля хвойных в насаждении и общая древесная фитомасса медленно изменяются на протяжении десятилетий. Сукцессионные переходы такого типа можно рассматривать как аналоги фазовых переходов второго рода в физических системах.
Настоящая работа посвящена разработке моделей, описывающих сукцесссионные переходы в лесных сообществах как фазовые переходы первого и второго рода. При этом в модели экологических фазовых переходов будут рассматриваться только возможности перехода из одной фазы в другую, и не будет рассматриваться «кинетика» этих переходов и даваться оценка времени этих фазовых переходов.
Смена травянистого ценоза на древесный как примере фазового перехода первого рода
Будем полагать, что особи некоторого вида с массой (численностью) Х(i) существуют в границах парцеллы и занимают некоторую часть S(i) от общей площади S ценоза. Будем также предполагать, что между особями данного вида существуют взаимодействия двух типов – кооперативное и конкурентное. В отсутствии других видов на территории при заданной численности Х(i) кооперативное взаимодействие будет способствовать «стягиванию» особей и уменьшению площади S(i). Конкурентное же взаимодействие будет приводить к «отталкиванию» особей и увеличению площади S(i) парцеллы. Площадь, занимаемая видом, не будет изменяться, когда интенсивности кооперативного и конкурентного взаимодействий будут равны.
Интенсивность конкурентного взаимодействия I1 для заданных видов будем полагать прямо пропорциональной плотности популяции (биомассы) и обратно пропорциональной площади территории, занимаемой видом. При этом необходимо учесть, что эта площадь не может быть бесконечно малой, так как в любом случае особи не могут размещаться в одной точке пространства. Введем для учета этого обстоятельства параметр b – минимальную площадь, на которой могут размещаться особи данного вида с численностью Х. Тогда
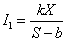
|
(1) |
где k – некоторая константа.
Кооперативное взаимодействие I2 будем полагать обратно пропорциональной некоторой степени величины площади парцеллы:
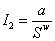 |
(2) |
В стационарном состоянии при отсутствии внешнего воздействия на особи в парцелле будет выполняться равенство I1 = I2 или
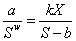
|
(3) |
Из (3) следует, что в равновесном состоянии связь между численностью популяции Х и площадью Sбудет выражаться уравнением
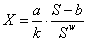
|
(4) |
Из (4) следует, что при отсутствии внешнего воздействия на вид возможно бесконечное число равновесных состояний вида с определенными значениями его численности (биомассы) и занимаемой площади, среди которых можно выделить состояние с максимальной при данных значениях параметров уравнений взаимодействий численностью популяции. В частности, при w=2 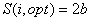 . .
Однако при анализе необходимо учесть воздействие на данный вид других видов, способное привести к уменьшению площади, занимаемой этим видом, и даже к вытеснению его из сообщества. Фактически внешнее конкурентное воздействие будет действовать точно так же, как внутривидовое кооперативное взаимодействие особей в парцелле, стремясь «стягивать» особи в парцелле с уменьшающейся площадью. Равновесное состояние популяции в пределах такой парцеллы при заданном внешнем воздействии на i-ый вид р(i) (экологическом давлении) будет выражаться уравнением
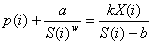
|
(5) |
При w=2 уравнение (5) можно рассматривать как экологический аналог уравнения Ван дер Вальса (Дмитриев, 2004).
При заданном значении численности Х(i) вида связь между внешним экологическим давлением р(i) и площадью S(i), занимаемой этим видом, отражена на рисунке 1.
Как видно из рисунка 1, форма кривых р(S) зависит от значения численности популяции Х. При значениях X > X0 кривая р(S) монотонна. При некотором значении X = X0 кривая р(S) имеет перегиб, при значениях X < X0 кривая р(S) немонотонна.
Предположим, что интенсивность внешнего экологического давления на популяцию, находящуюся в данной парцелле, сначала было велико, а затем стало уменьшаться. Если численность популяции Х>X0, то с уменьшением внешнего экологического давления площадь, занимаемая данной популяцией в стационарных условиях, будет монотонно возрастать. Напротив, если при малом начальном экологическом давлении оно начинает возрастать, то площадь парцеллы, занимаемой данной популяцией в стационарных условиях, начинает монотонно уменьшаться. Если же численность популяции X < X0, то, начиная с определенного значения р, увеличения площади парцеллы, занимаемой данной популяцией, можно добиться, только увеличивая, а не уменьшая внешнее экологическое давление. Происходит это из-за немонотонного поведения кривой р(S) (рис. 1). Обратно, при увеличении внешнего экологического давления, сопровождаемого уменьшением площади парцеллы, занимаемой популяцией, начиная с некоторой величины площади для дальнейшего ее уменьшения нужно будет не увеличивать, а уменьшать внешнее экологическое давление. Такое поведение модельной системы представляется невозможным в реальных условиях. Однако этого парадокса можно избежать, если предположить, что на критическом участке (1,2) кривой р(S) реальное изменение переменных р и S происходит так, как это показано на рисунке 1 (прямая АВ). В этом случае X < X0 и изменение площади парцеллы, занимаемой популяцией, на участке (1,2) будет происходить при постоянном значении внешнего экологического давления р. После достижения границ этого участка вновь будет иметь место монотонная обратная связь между величиной внешнего экологического давления и площадью территории, занимаемой популяцией. В этом случае при постоянном значении р будет иметь место скачок величины площади, занимаемой изучаемым видом. В физических системах такое поведение соответствует фазовому переходу первого рода.
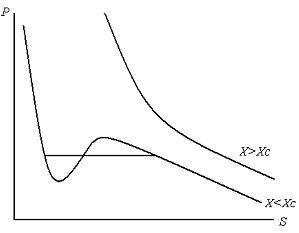
Рисунок 1 – Cвязь между внешним экологическим давлением р и площадью S, занимаемой видом
Рассмотрим использование модели для описания взаимодействий «лес - травянистые растения». Будем рассматривать сформировавшийся лесной ценоз общей площадью Sm, в котором травянистые растения с фитомассой Х занимают площадь Sg. Ограничение площади, занимаемой травянистыми растениями, происходит за счет внешнего экологического давления р со стороны древесной растительности. Пусть в некоторый момент времени древесные растения частично или полностью погибнут. В терминах вышеизложенной модели это означает, что внешнее экологическое давление упало (возможно, до нуля). В этом случае, если фитомасса травянистой растительности велика (больше критического значения Х0) будет иметь место замещение лесного ценоза травянистым и увеличение площади, занимаемой травянистыми растениями. Если же фитомасса травянистых растений до деградации древесной растительности была мала, то в случае, когда внешнее экологическое давление на травянистые растения уменьшится до значения р0, произойдет резкий скачок величины площади, занимаемой травой. Подобного рода явления характерны для ситуаций после «быстрых» критических явлений в экосистемах, таких, как пожары и ветровалы.
Смена пород в лесном ценозе как пример фазового перехода второго рода
Наряду с фазовыми переходами первого рода можно рассматривать и фазовые переходы второго рода, когда некоторая характеристика (параметр порядка) в насаждении меняется непрерывно. Модель фазового перехода второго рода будем использовать для описания сукцессионного перехода «лиственное насаждение – хвойное насаждение».
Введем в качестве параметра порядка величину q (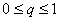 ), характеризующую долю деревьев хвойных пород в насаждении. Значение параметра порядка q = 1 соответствует чистым хвойным насаждениям, значение q = 0 – чистым лиственным насаждениям, промежуточные значения q характеризуют смешанное насаждение. Сукцессионный переход «лиственные – хвойные» будет начинаться, когда в лиственном насаждении появятся хвойные деревья и параметр порядка q станет отличным от нуля. Введем также понятие потенциала экологического риска G. Величина G характеризует вероятность трансформации (сукцессионного перехода) ценоза. Чем больше значение G, тем больше вероятность трансформации. Конечно, записать в общем виде уравнение зависимости G от разнообразных параметров, характеризующих ценоз и воздействие на него внешних факторов, весьма затруднительно. Тем не менее, независимо от конкретной формы уравнения для G, можно ввести оптимизационный принцип, регулирующий процесс сукцессионного фазового перехода, как условие ), характеризующую долю деревьев хвойных пород в насаждении. Значение параметра порядка q = 1 соответствует чистым хвойным насаждениям, значение q = 0 – чистым лиственным насаждениям, промежуточные значения q характеризуют смешанное насаждение. Сукцессионный переход «лиственные – хвойные» будет начинаться, когда в лиственном насаждении появятся хвойные деревья и параметр порядка q станет отличным от нуля. Введем также понятие потенциала экологического риска G. Величина G характеризует вероятность трансформации (сукцессионного перехода) ценоза. Чем больше значение G, тем больше вероятность трансформации. Конечно, записать в общем виде уравнение зависимости G от разнообразных параметров, характеризующих ценоз и воздействие на него внешних факторов, весьма затруднительно. Тем не менее, независимо от конкретной формы уравнения для G, можно ввести оптимизационный принцип, регулирующий процесс сукцессионного фазового перехода, как условие 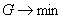 . Использование такого оптимизационного принципа означает, что система (ценоз) стремится к достижению состояния, при котором экологический риск трансформации минимален. . Использование такого оптимизационного принципа означает, что система (ценоз) стремится к достижению состояния, при котором экологический риск трансформации минимален.
Вблизи точки фазового перехода второго рода число факторов, влияющих на поведение системы, обычно сильно уменьшается и функция G зависит только от параметра порядка (Брус, Каули, 1984; Ландау, Лифшиц, 1964). Зависимость 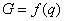 функции экологического риска от параметра порядка можно представить степенным рядом, в котором опущены нечетные члены ряда разложения G (в противном случае имел бы место скачкоообразный фазовый переход первого рода), и четные члены ряда со степенями, большими четырех (ввиду их малости по сравнению с первыми членами разложения функции функции экологического риска от параметра порядка можно представить степенным рядом, в котором опущены нечетные члены ряда разложения G (в противном случае имел бы место скачкоообразный фазовый переход первого рода), и четные члены ряда со степенями, большими четырех (ввиду их малости по сравнению с первыми членами разложения функции 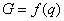 ): ):
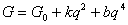 |
(6) |
где G0, k, b - коэффициенты.
В качестве фактора, оказывающем влияние на возможность фазового перехода от лиственного насаждения к хвойному, введем величину фитомассы Х лиственной части насаждения. Воздействие такого фактора можно ввести в форме линейной зависимости от Х коэффициента k при квадратичном члене уравнения (1): 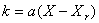 . Тогда из (6) имеем: . Тогда из (6) имеем:
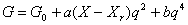
|
(7) |
где а, b – постоянные коэффициенты, X - фитомасса лиственной части насаждения, Xr - критическое значение фитомассы лиственной части насаждения, при достижении которого в насаждении появляются хвойные деревья и начинается сукцессионный фазовый переход.
Исходя из оптимизационного принципа, устойчивому состоянию ценоза должно соответствовать минимальное значение G. Условие минимума функции (7) запишется стандартным образом:
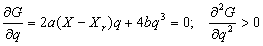
|
(8) |
Уравнение (8) имеет два решения:
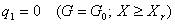
|
(9) |
и
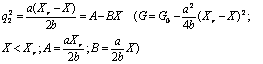
|
(9) |
Так как согласно модели при Х = 0 q2 = 1, то в (10) коэффициент А =1.
Устойчивое решение q = 0 соответствует случаю, когда запас лиственных насаждений больше некоторого критического значения Хr. Второе решение реализуется, когда запас лиственного насаждения становится меньшим Хr и оно начинает трансформироваться в смешанное. Согласно уравнениям (9) и (10), значение функции экологического риска G для смешанного насаждения меньше, чем для лиственного насаждения (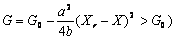 , то есть хвойные насаждения более устойчивы. Для смешанных насаждений, согласно (10), существует линейная связь между квадратом параметра порядка q2 и запасом X лиственной части насаждения. Когда сукцессионный переход от лиственных к хвойным закончился и запас лиственной части насаждения становится равной нулю, то q2 = 1. , то есть хвойные насаждения более устойчивы. Для смешанных насаждений, согласно (10), существует линейная связь между квадратом параметра порядка q2 и запасом X лиственной части насаждения. Когда сукцессионный переход от лиственных к хвойным закончился и запас лиственной части насаждения становится равной нулю, то q2 = 1.
Для верификации модели необходимо, используя методы регрессионного анализа, по величине коэффициента детерминации R2 определить степень согласия натурных данных с линейной моделью (10) и найти коэффициенты линейного регрессионного уравнения связи между Х и q2. Величину критического запаса Xr (10) можно вычислить через параметры уравнения (10) по формуле 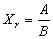 . .
Таким образом, в модели сукцессионного фазового перехода второго рода связь между фитомассой лиственной части насаждения и квадратом параметра порядка выражается двумя прямыми с изломом в точке {Xr, 0} (рис. 2).
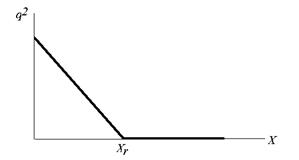
Рисунок 2 – Теоретическая связь между параметром порядка q2 и фитомассой X лиственной части насаждения
Согласно модели фазовых переходов, если фитомасса лиственных в насаждении превосходит критическое значение Xr, в таких насаждениях не будут встречаться деревья хвойных пород. При уменьшении фитомассы лиственных ниже критического значения в насаждении должны появиться хвойные деревья, а связь между квадратом параметра порядка насаждения и фитомассой лиственных в этом насаждении должна быть линейной.
Для верификации модели были привлечены таксационные данные лесоустройства Таловского и Предивинского лесничеств Большемуртинского опытно-показательного лесхоза, выполненного с использованием программно-методических материалов и схем типов леса, составленных Институтом леса СО РАН (Региональные…, 2007). Объектами исследований служили коренные смешанные темнохвойные и производные хвойно-лиственные и лиственные леса северного макросклона Идарского белогорья Восточного Саяна и леса Средней Сибири в пределах подзоны южной тайги и южной части Енисейского кряжа. Исходная информация собиралась в течение почти 30 лет при маршрутных исследованиях с закладкой временных пробных площадей и профилей, а также наблюдений экспериментальных посадок, постоянных объектов с рубками ухода, и другими работами. Эта информация позволила оценить масштаб смены хвойных пород лиственными и представить общие сценарии восстановительных смен лиственных пород хвойными.
В анализ включены данные для 3269 таксационных выделов в Предивинском лесничестве и для 2070 выделов – в Таловском, относящиеся к наиболее распространенным типами леса и их сериям (табл. 1).
Таблица 1 - Параметры уравнений сукцессионного перехода от лиственных к темнохвойным насаждениям
Серии типов темнохвойных и лиственных лесов |
Число
выделов |
Статистические показатели |
R2 |
В |
А |
Xr=A/B |
Предивинское лесничество |
Вейниково-крупнотравная |
246 |
0,757 |
0,005 |
0,883 |
173,2 |
Осочково-зеленомошная |
123 |
0,847 |
0,005 |
0,899 |
172,9 |
Вейниково-осочково-зеленомошная |
332 |
0,897 |
0,006 |
0,956 |
162,0 |
Мелкотравно-вейниковая |
388 |
0,805 |
0,006 |
0,910 |
154,2 |
Вейниковая |
565 |
0,833 |
0,006 |
0,908 |
146,4 |
Хвощево-вейниковая |
166 |
0,749 |
0,006 |
0,875 |
138,9 |
Разнотравно-осочковая |
839 |
0,790 |
0,006 |
0,875 |
136,7 |
Злаково-крупнотравная |
552 |
0,838 |
0,007 |
0,968 |
132,6 |
Кустарниково-разнотравная |
58 |
0,920 |
0,011 |
1,001 |
91,0 |
Таловское лесничество |
Вейниково-осочково-зеленомошная |
125 |
0,910 |
0,005 |
0,966 |
205,5 |
Вейниково-мелкотравно-осочковая |
345 |
0,860 |
0,005 |
0,952 |
198,4 |
Хвощево-зеленомошная |
213 |
0,922 |
0,005 |
0,976 |
187,6 |
Мелкотравно-зеленомошная |
270 |
0,923 |
0,005 |
0,975 |
180,6 |
Вейниково-папоротниковая |
35 |
0,961 |
0,006 |
0,980 |
169,0 |
Осочково-разнотравная |
248 |
0,549 |
0,005 |
0,775 |
164,8 |
Злаково-крупнотравная |
70 |
0,842 |
0,006 |
0,920 |
164,3 |
Вейниково-крупнотравная |
323 |
0,790 |
0,005 |
0,863 |
159,7 |
Мелкотравно-вейниковая |
114 |
0,741 |
0,007 |
0,966 |
142,0 |
Разнотравно-осочково-зеленомошная |
139 |
0,820 |
0,007 |
0,953 |
140,1 |
Разнотравно-брусничная |
22 |
0,814 |
0,007 |
0,927 |
136,3 |
Разнотравная |
166 |
0,828 |
0,077 |
0,893 |
115,9 |
На рисунке 3 показано изменение породного состава насаждений по 20-летним классам возраста (1251 выдел в границах Таловского лесничества) в ходе сукцессионных переходов в разнотравные серии типов леса. Эта серия типов леса была выбрана для анализа как типичная, отражающая сукцессии в подзоне южной тайги. Для каждого типа леса суммировались доли пород в составе насаждения и вычислялась доля данной породы в каждом возрастном классе. Запас каждого возрастного класса вычислялся как среднее значение для выделов.
Анализ показал, что сукцессионный переход от лиственных к хвойным насаждениям выражается в сопряженном изменении породного состава насаждения и запаса древесины в нем (рис. 3 и 4).
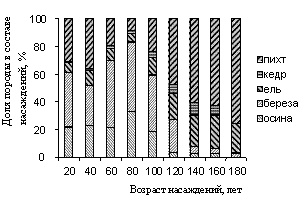
Рисунок 3 – Возрастные изменение породного состава насаждений (Таловское лесничество Большемуртинского лесхоза Красноярского края)

Рисунок 4 – Динамика запаса древесины разных пород в насаждениях (Таловское лесничество Большемуртинского лесхоза Красноярского края)
На начальном этапе сукцессии (в возрасте деревьев до 20 лет) древесный ценоз представляют собой смешанное насаждение, где доля лиственных (березы и осины) составляет около 60%. В течение следующих 30-40 лет в этих насаждениях происходит сукцессионный переход, в результате которого появляются насаждения, в которых доля лиственных превосходит 80%. Следующий сукцессионный переход начинается, когда возраст лиственных в насаждении достигает 70-80 лет, и длится около 100-150 лет. За это время лиственный лес заменяется на темнохвойный с преобладанием (свыше 90% деревьев) пихты и ели. При этом к 180 годам осина полностью элиминируется из состава насаждения.
В ходе сукцессии изменяется и запас древесины в насаждениях (рис. 4). В возрасте около 60 лет запас древесины лиственных составлял примерно 75% от общего, достигая своего максимума к 80-100-летнему возрасту, а к 180 годам снижается до 15% от общего. По достижению возраста 260 лет основной вклад (около 90%) в запас древесины насаждений вносят хвойные.
Согласно уравнению (10), если модель сукцессионных фазовых переходов второго рода корректна, то должна существовать линейная связь между квадратом параметра порядка насаждения и запасом лиственных в смешанном насаждении. Используя данные по отдельным выделам и пробным площадям, были вычислены величины q2 и изучена связь q2 с запасом Х лиственной части насаждений на этих объектах (рис. 5).
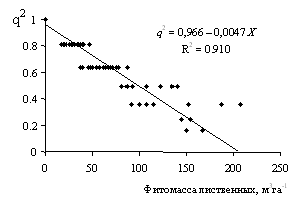 Рисунок 5 - Связь между величинами квадрата параметра порядка и запасом лиственной части смешанных насаждений (выделы мелкотравно-осочково-зеленомошной серии типов леса в Таловском лесничестве Большемуртинского лесхоза, критическое расчетное значение Xr = 205 м3га-1) Рисунок 5 - Связь между величинами квадрата параметра порядка и запасом лиственной части смешанных насаждений (выделы мелкотравно-осочково-зеленомошной серии типов леса в Таловском лесничестве Большемуртинского лесхоза, критическое расчетное значение Xr = 205 м3га-1)
Анализ этих данных показал, что для смешанных насаждений прогнозируемая в модели фазовых переходов связь между квадратом параметра порядка и запасом лиственной части, отраженная на рисунке 5, хорошо согласуется теоретическим графиком на рисунке 2. Как видно из рисунка 5, связь между q2 и запасом лиственной части смешанных насаждений хорошо описывается линейным регрессионным уравнением. Величина коэффициента детерминации R2 = 0,91 близка к теоретическому значению R2 = 1 для линейных уравнений, а свободный член регрессионного уравнения А = 0,966 весьма близок к теоретическому значению А = 1. Имеющиеся данные по объектам мелкотравно-осочково-зеленомошной серии типов леса в Таловском лесничестве не характеризуют достаточно редкие в этой серии лиственные насаждения, поэтому предсказываемого теоретически излома прямой q2 = A – BX на рисунке 5 не наблюдается.
Для вейниково-крупнотравных типов леса, где присутствуют как смешанные, так и лиственные насаждения при различных значениях запаса лиственных пород величина параметра порядка q насаждения близка или равна нулю (рис. 6). Этот результат хорошо согласуется с моделью на рисунке 2.
Насколько близка к линейной (теоретической) связь между квадратом параметра порядка насаждения и фитомассой его лиственной части? В таблице 1 (нижняя часть) приведены параметры
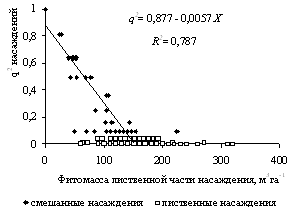
Рисунок 6 - Связь между величинами параметра порядка q и запасом Х лиственной части насаждений для смешанных и лиственных насаждений вейниково-крупнотравной серии типов леса (критическое расчетное значение Xr = 150 м3га-1)
регрессионных уравнений связи между q2 и запасом Х лиственной части смешанных насаждений в Таловском лесничестве Большемуртинского лесхоза. Как видно из таблице 1, связь между квадратом параметра порядка и запасом лиственной части смешанного или хвойного насаждения для всех серий и типов леса линейна. Коэффициенты детерминации R2 и значения свободного члена регрессионных уравнений в большинстве случае превосходят 0,9. Это говорит о высокой степени согласия модели фазовых сукцессионных переходов и натурных данных. Однако критические значения запаса лиственного насаждения, когда в нем появляются хвойные деревья, для различных типов и серий типов леса изменяются в широких пределах – от 205 м3га-1 в разнотравно-осочково-зеленомошной серии типов леса до величины 95 м3га-1 для осоково-сфагновой (табл. 1). Поэтому нет оснований утверждать, что критическое значение фитомассы при сукцессионном переходе есть константа.
Используя имеющиеся данные, были вычислены параметры уравнений (10) и для насаждений на территории Предивинского лесничества Большемуртинского лесхоза (табл. 1, верхняя часть). И в этом случае близость величин коэффициентов детерминации R2 и свободных членов регрессионных уравнений к 1 говорят о том, что натурные данные хорошо согласуются с теоретической моделью.
Анализ данных для насаждений одной серии типов леса, но разных бонитетов показал, что критическое значение запаса лиственных монотонно уменьшается от насаждений первого бонитета до насаждений третьего бонитета (табл. 2).
Таким образом, критическая фитомасса лиственных, при которой в таежных лесах Средней Сибири происходит переход от лиственных к хвойным насаждениям, зависит как от типа леса, так и от его бонитета. Существование такой зависимости объясняет появление на рисунке 6 точек, характеризующих лиственные насаждения с q = 0, но расположенных левее критической точки Xr = 150 м3га-1. Как показывает более детальный анализ, эти точки характеризуют низкобонитетные насаждения.
Таблица 2 - Параметры уравнений сукцессионного перехода от лиственных к темнохвойным насаждениям разного бонитета
Серия типов леса |
Бонитет |
R2 |
B |
A |
Хr=A/B |
Вейниково-крупнотравная |
3 |
0,730 |
0,0054 |
0,891 |
165,0 |
4 |
0,866 |
0,0048 |
0,862 |
179,5 |
Вейниковая |
2 |
0,775 |
0,0052 |
0,816 |
156,0 |
3 |
0,812 |
0,0061 |
0,862 |
141,3 |
4 |
0,888 |
0,0111 |
1,023 |
92,2 |
Разнотравно-осочковая |
1 |
0,856 |
0,006 |
0,910 |
156,9 |
2 |
0,730 |
0,006 |
0,797 |
135,1 |
3 |
0,840 |
0,007 |
0,910 |
128,2 |
Безусловно, существуют определенные отклонения натурных данных от расчетов по предложенной модели. Эти отклонения могут быть следствием целого ряда причин. Например, в модели не учитывается возможность существования в молодых насаждениях отдельных крупномерных деревьев, сохранившихся от предыдущих поколений. Вклад таких деревьев в величину параметра порядка будет мал, но фитомасса таких деревьев может быть весьма большой, что приведет к смещению точки на плоскости 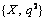 , характеризующей данное насаждения относительно теоретической прямой. , характеризующей данное насаждения относительно теоретической прямой.
Заключение
Анализ и сопоставление модели сукцессионных фазовых переходов с натурными данными показали, что имеет место достаточно высокий уровень согласия между моделью и натурными лесотаксационными данными. Модель позволяет объяснить направленность сукцессионных процессов и определить, на каком этапе развития экологических процессов следует ожидать сукцессионных переходов. Анализ данных натурных наблюдений с использованием предлагаемого подхода позволяет для конкретных регионов и определенных типов леса вычислить критические значения фитомассы насаждения, при достижении которых начинается сукцессионный переход. Знание этих величин важно при средне- и долгосрочных прогнозах лесообразовательного процесса.
Ранее для описания сукцессионных переходов и роста фитомассы видов в растительных сообществах была предложена модель с двумя устойчивыми состояниями (Исаев и др., 2004, 2005). Одно из этих состояний характеризует величину фитомассы травянистого сообщества, другое – величину фитомассы древесного насаждения. Переход из состояния с преобладанием травянистой растительности в состояние с преобладанием древесных растений происходит, когда фитомасса насаждения превысит
некоторое критическое значение. Модель экологических фазовых переходов в значительной степени схожа с феноменологической моделью роста лесных насаждений. Существенное различие, однако, состоит в том, что по модели фазовых переходов возможно не только обосновать существование критической точки изменения характеристик лесного ценоза, но рассчитать величину критической биомассы, при достижении которой происходит экологический фазовый переход.
Библиографический список
- Александрова, В.Д. Изучение смен растительного покрова / В.Д. Александрова // Полевая геоботаник М.-Л.: Наука, 1964. - Т.3. - С. 300-447.
- Брус, А. Структурные фазовые переходы / А. Брус, Р. Каули,. М.: Мир, 1984. – 408 с.
- Дмитриев, А.В. Основы статистической физики материалов / А.В. Дмитриев. М.: Изд-во МГУ, Наука, 2004. – 668 с.
- Замолодчиков, Д.Г Углерод в лесном фонде и сельскохозяйственных угодьях России / Д.Г. Замолодчиков, и др. - М.: Товарищество научных изданий КМК, 2005. – 200 с.
- Исаев, А.С. Феноменологическая модель лесообразовательного процесса / А.С. Исаев, В.Г. Суховольский, Р.Г. Хлебопрос // ДАН, 2004. - т. 398. - № 6. - С. 845 – 848.
- Исаев, А.С Моделирование лесообразовательного процесса: феноменологический подход / А.С. Исаев, В.Г. Суховольский, Р.Г. Хлебопрос // Лесоведение, 2005. - № 1. - С. 1 -9.
- Ландау, Л.Д. Cтатистическая физика / Л.Д. Ландау, Е.М. Лифшиц. – М.: Наука, 1964. – 567 с.
- Миркин, Р.М. Фитоценология. Принципы и методы / Р.М. Миркин, Г.С. Розенберг. – М.: Наука, 1978. – 212 с.
- Региональные проблемы экосистемного лесоводства /под ред. А.А. Онучина / - Красноярск: Институт леса им. В.Н.Сукачева СО РАН, 2007. – 330 с.
- Сукачев, В.Н. Динамика лесных биогеоценозов / В.Н. Сукачев // Основы лесной биогеоценологии. Под ред. В.Н. Сукачева и Н.В. Дылиса. – М.: Наука, 1964. – С. 458-486.
- Soukhovolsky, V.G. Models of phase transition in forest ecosystems / V.G. Soukhovolsky // ECEM’07, Conference proceedings. Trieste, Italy, 2007. - p. 505 – 506.
|


