|
"Хвойные бореальной зоны" 2007г.,№4-5, с. 467- 473
Интенсивность гетеротрофного дыхания в сосняках средней тайги: сравнительный анализ методов оценки
Трефилова О.В.
Институт леса им. В.Н. Сукачева СО РАН
660036 Красноярск, Академгородок, 50; е-mail: trefilova@ksc.krasn.ru
Интенсивность гетеротрофного дыхания (Rh) оценивали с помощью двух различных подходов: метода «изоляции» и метода «интеграции компонентов». В ходе экспериментальных исследований в молодняках и перестойных насаждениях зеленомошного и лишайникового типов леса оба используемых метода показали удовлетворительную сходимость результатов. Метод «интеграции компонентов» предпочтительнее использовать для оценки интенсивности гетеротрофного дыхания в экосистемах с низким запасом (или при отсутствии) грубых древесных остатков. В среднем интенсивность Rh почв зеленомошных сосняков составляет 27 ц С га-1год-1, в лишайниковых – 18 27 ц С га-1год-1. Основная масса углерода в потоке Rh формируется при разложении подстилки и корневого детрита, относительное участие грубых древесных остатков составляет 14-15% в сосняках лишайниковой и 7-10% в сосняках зеленомошной группы леса.
Intensity heterotrophic respiration (Rh) estimated by two various approaches: a "isolation" method and a "integration of components» method. During experimental researches in saplings and over-mature pine forests with larch or green moss methods have shown satisfactory convergence of results. The method of "integration of components» is more preferable for using to intensity Rh estimation low storage the coarse woody debris ecosystem. In pine forest with green moss intensity heterotrophic soil respiration averages 2.7 t ha-1year-1, in pine forests with lichen - 18 t ha-1year-1. The main part of Rh carbon is formed at decomposition of a forest litter and dead root mass, relative participation of the coarse woody debris makes 14-15 % in pine forests with lichen and 7-10 % in pine forests with green moss.
Введение
Гетеротрофное дыхание (Rh), как и чистая первичная продукция (NPP), являются интегральными показателями, определяющими статус экосистемы в биосфере: «сток» или «источник» для углерода атмосферы. Величина NPP служит приходной статьёй баланса углерода (С), характеризует продуцирование атмосферного углерода в тканях растений в процессе фотосинтетической ассимиляции, а Rh - расходная статья баланса, показывающая, какое количество С ежегодно возвращается в атмосферу, освобождаясь в процессах деструкции растительных остатков и мобильной части гумуса почв.
Экспериментальные оценки гетеротрофного дыхания лесных экосистем в большинстве своем получены на основе двух различных подходов. Первый – метод «изоляции» - базируется на определении интенсивности и удельной скорости разложения фиксированной массы растительных остатков на поверхности и в толще почвы (Титлянова, 1977; Ведрова, 1997, 2005 Vedrova и др. 2006). Гетеротрофное дыхание рассматривается как интенсивность минерализации легкоминерализуемой фракции органического вещества почвы, т.е. как сумма минерализации лесной подстилки (АОСО2), корневого детрита (RDCO2), грубых древесных остатков (CWDCO2), свежеобразованных гумусовых веществ (NhumCO2), и подвижных продуктов собственно гумуса почвы (ShumCO2) (Ведрова и др., 2002; Ведрова, 2005; Vedrova et.al., 2006). Часть новообразованного гумуса аккумулируется в почве (Nhumaccum), замещая минерализовавшуюся
_______________________
*Исследования выполнялись при поддержке РФФИ (06-04-90596) и проекта МНТЦ 2770
часть подвижного гумуса. Таким образом, суммарный поток углерода в атмосферу при разложении органического вещества почвы рассчитывается как:
Rh = АОСО2 + RDCO2 + CWDCO2 + NhumCO2 + (ShumCO2 - Nhumaccum) (1)
Основным недостатком метода является необходимость проведения длительных натурных экспериментов для получения адекватных результатов. Неопределенности связаны с рядом методических сложностей. Поскольку интенсивность разложения определяется по потере массы образцов за время эксперимента, растительный материал разлагается в сетчатых мешочках, диаметр ячей которых может ограничивать доступ деструкторов более крупного размера. При малой длительности эксперимента нарушаются условия трансформации образцов: изменяются воздушный и водный режим. Проведение экспериментов в полевых условиях осложняется проникновением живых тонких корней во внутреннее пространство образца, что искажает реальные показатели скорости разложения. В результате наблюдается значительная вариабельность получаемых данных, особенно при разложении образцов в слое подстилок (Hanson et.al, 2000).
Неопределенности метода могут быть связаны и с определением интенсивности гумификации в процессе разложения растительных остатков. Она оценивается разницей количества углерода соединений, извлекаемых из образцов слабым раствором гидроксида натрия в начале и при окончании эксперимента, или по выносу водорастворимого углерода с лизимитрическими растворами (Кауричев и др., 1989, Чагина, 1976, 1970, Ведрова, 1997). Получаемые оценки в целом не противоречат данным более тонкого и точного определения интенсивности новообразования гумусовых веществ с применением методов радиоуглеродного датирования (Фокин, 1974, 1978; Ганжара, 1979, Багаутдинов, Куватов, 1993), проведение которого требует специального оборудования и крайне затруднительно в естественных условиях.
Второй подход – метод «интеграции компонентов» - основан на положении о том, что поступающий в атмосферу углекислый газ является результатом дыхания корней живых растений и разложения почвенной биотой мёртвых органических остатков и гумуса (Мина, 1962; Кайбияйнен и др., 1999; Ларионова и др., 2003, Благодатский, 1993 и др.). Использование этого метода позволяет разделить гетеротрофные и автотрофные потоки СО2: измерения проводят на поверхности ненарушенной почвы, содержащей корни растений, и на подготовленных участках почвы, лишенных корней (Цельникер, 2005). Этот метод также не лишён перечисленных выше недостатков: нарушается естественное сложение почвы, а значит, и условия, влияющие на скорость минерализации. Почва обогащается кислородом. При извлечении растений угнетается активность микроорганизмов ризосферы, а соответственно и вклад их дыхания в общий поток СО2 (Hanson et al, 2000; Благодатский и др., 1993). Ограничением использования метода является также необходимость организации постоянных наблюдений из-за суточной, месячной, сезонной динамики интенсивности выделения СО2, обусловленной изменениями температуры и влажности среды, периодичностью поступления отмирающих органов растений (Макаров, 1958; Наумов, 1994). Чтобы учесть в лесных экосистемах ярко выраженную пестроту, в том числе пространственную неоднородность растительного покрова, запаса и состава мёртвой растительной органики, корней живых растений, необходимо увеличение числа замеров концентрации углекислого газа в приземном слое воздуха.
Для измерения потока СО2 из почвы разработано
множество методов, различающихся способом отбора (микрометеорологический, профильный, камерный) и техникой анализа проб (абсорбционный, газохроматографический или с помощью инфракрасного анализатора и др.) (Шарков, 1984, 1987 а, б; Макаров, 1975, 1988; Ларионова, Розонова, Самойлов, 1988; и др.). Литературные данные относительно надёжности методов противоречивы. По данным (Ларионова, Иванникова, Демкина, 1993), применение различных методов на одном и том же объекте даёт близкие результаты. В то же время немало работ, выводы которых указывают на несовпадение данных, полученных разными методами (Мина, 1962; Курганова, Кудеяров, 1998; Freijer, Bouten, 1991 и др.).
Несмотря на недостатки, оба метода определения гетеротрофного потока углерода в равной степени популярны для исследований как локального, так и регионального уровня (Ведрова, 2005; Shvidenko, Nilsson, 2003; Кудеяров, 2005; Кудеяров, Курганова, 2005 и др.). Выбор остается за исследователем и в большей степени определяется материально-техническими возможностями экспериментальной базы.
Целью настоящей статьи является сравнительная оценка вышеприведенных методов определения интенсивности гетеротрофного дыхания почвы лесных экосистем.
Объект и методика
Исследования проводили на четырёх постоянных пробных площадях в сосняках междуречья Тугулана и Дубчеса, в пределах Тугуланской котловины Енисейской равнины (60053?с.ш., 89038? в.д.). Постоянные пробные площади характеризуют сосновые древостои двух типов леса: сосняки зеленомошные (С. зм.) в возрасте 20 и 250 лет и лишайниковые (С. лш.) – 15 и 260 лет. Все насаждения пройдены пожарами. Таксационная характеристика древостоев приведена в таблице 1.
Таблица 1 - Основные таксационные характеристики фитоценозов пробных площадей
№ |
Состав |
Возраст, лет |
Средние |
Густота,
шт га-1 |
Сумма площадей поперечного сечения, м2 га-1 |
Запас,
м3 га-1 |
диаметр, см |
высота, м |
Зеленомошная группа типов леса |
1 |
7С 3Б |
С |
20 |
7,0 |
7,3 |
4020 |
15,0 |
70,8 |
Б |
23 |
7,5 |
8,9 |
1360 |
6,0 |
53,5 |
Ос |
12 |
6,0 |
8,1 |
40 |
0,1 |
1,0 |
2 |
10С |
С |
250 |
25,4 |
20,8 |
439 |
30,4 |
368,9 |
С |
90 |
12,1 |
13,3 |
182 |
2,1 |
16,2 |
Б |
- |
3,0 |
4,5 |
102 |
0,1 |
0,4 |
Ос |
- |
2,6 |
3,8 |
187 |
0,1 |
0,3 |
Лишайниковая группа типов леса |
3 |
10С |
С |
15 |
3,4 |
3,4 |
9237 |
8,1 |
25,7 |
4 |
10С |
С |
260 |
35,7 |
22,1 |
283 |
28,5 |
366,3 |
С |
100 |
10,3 |
11,0 |
86 |
0,7 |
5,1 |
Примечание: С - сосна; Б - берёза; Ос – осина.
Рельеф территории характеризуется чередованием уплощённых холмов, валов, гряд. Сосняки ксерофильной группы, как правило, развиваются на иллювиально-железистых подзолах, в верхней части пологих склонов и на водоразделах. Мезофильные сосняки приурочены к западинам и нижним частям склонов. Переувлажнение почв зеленомошных сосняков обусловливает формирование иллювиально-железистого глубокооглеенного подзола.
Климат исследуемой территории – континентальный, с абсолютной амплитудой температур 90- 950С, при среднегодовых показателях равных – 4,7- 4,50С. Годовое количество осадков составляет 400-650 мм. ГТК изменяется в пределах от 1,2 до 1,6 (Куваев и др., 2001).
В рамках первого подхода (метод «изоляции») закладывался полевой эксперимента по разложению растительных остатков на поверхности и в слое почвы 0-5 см. Исходным материалом для него служили фракции растительного материала, полученные при учете запаса опада, подстилки (АО), грубых древесных остатков (CWD) и корневого детрита (RMM) на пробных площадях (Трефилова, 2006). Для проведения полевого эксперимента использовались образцы подстилки (n = 5) и корневого материала (n = 5), составленные с учётом среднего запаса мертвых органических остатков на поверхности и в толще почвы. Предварительно взвешенный растительный материал помещался в капроновые мешочки с размером ячеи 2 мм. Мешочки с подстилкой закладывали в ферментативный слой, образцы корневого материала - в минеральный слой почвы, на глубину 0-5 см.
Спустя год образцы вынимались, определялась их масса, влажность. В исходных и изъятых образцах определялась концентрация углерода в экстрактах, полученных при последовательной обработке образцов дистиллированной водой и 0,1н раствором NaOH, в щелочной вытяжке определялась концентрация СГК (Пономарева, Плотникова, 1975).
Таким же способом анализировали образцы почвы для оценки концентрации в них углерода подвижной части гумуса. Содержание общего углерода в растительных образцах определяли методом Анстета в модификации Пономарёвой-Николаевой (Пономарева, Плотникова, 1975).
Интенсивность разложения рассчитывали по потере растительной массы за год. Поскольку при разложении растительного материала принимается, что его распад описывается экспонентой, для характеристики скорости разложения использовали величину константы разложения (Jenkinson, Rayner, 1977):
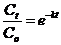 , ,
отсюда k = ln (Сt / С0) / t, (2)
где Сt и С0 масса образца в начале и конце эксперимента, t - время эксперимента, год; k – константа разложения с размерностью, обратной времени, год-1).
В качестве базовых показателей для расчёта скорости разложения грубых древесных остатков использовалась плотность здоровой (?0) и мёртвой древесины (?t): -k = ln (?t / ?0) / t. Период, на протяжении которого разлагается изучаемый фрагмент древесины (t), оценивался нами на основе косвенных показателей (давность лесохозяйственных мероприятий, ветровала, пожара, возраст самосева на пнях и брёвнах и т.п.). Результаты определения плотности древесины сосны по классам разложения в условиях Тугулан-Дубческого междуречья обсуждались нами ранее (Трефилова, 2006).
Разницу в содержании СГК в исходных и отобранных при окончании эксперимента образцах принимали за меру гумификации (Nhum) и использовали для расчета интенсивности гумификации (С, ц га-1год-1) (Ведрова, 2002).
Для расчета интенсивности закрепления почвой новообразованных гумусовых веществ (Nhumaccum) и минерализации подвижных продуктов гумуса почвы использовались экспериментальные оценки (Ведрова, 2005): соответственно, Nhumaccum составляет 48% «новых» гумусовых веществ, SHumCO2 – 4,6% запаса подвижного гумуса почвы.
Гетеротрофное дыхание (Rh, минерализационный поток) рассчитывали по приведенному выше уравнению 1.
Измерение интенсивности выделения углекислоты с поверхности почвы и определение вклада автотрофных и гетеротрофных источников СО2 проводили во втором эксперименте (метод «интеграции компонентов»). На каждой пробной площади выбиралось по два относительно однородных участка. Первый из них оставался ненарушенным. На втором удаляли слой подстилки и напочвенного покрова. Поскольку напочвенная растительность и слой мёртвого органического вещества на поверхности почвы практически неразделимы, подстилку и напочвенный покров снимали совместно. Затем на участке с площадки 15х15 см в четырёхкратной повторности до глубины 30 см извлекались почвенные монолиты. В получившиеся выемки устанавливали пластиковые цилиндры диаметром 15 см. Извлечённый почвенный субстрат освобождался от живых корней и укладывался в цилиндры. Выемки засыпались оставшимся субстратом. Слой напочвенного покрова и подстилки возвращали на прежнее место. Учитывая нарушения, сопутствующие закладке эксперимента, первые измерения на площадках проводили, спустя год (Hanson et. al, 2000; Цельникер, 2005).
Интенсивность выделения СО2 измеряли абсорбционным методом, условия и ход эксперимента аналогичны, описанным Э.Ф. Ведровой (1997). Измерения сопровождали определением температуры и влажности почвы. Расчёт интенсивности годового потока эмиссии углекислого газа произведён для вегетационного периода, продолжительность которого для данной территории составляет 132 – 150 дней (Куваев и др., 2001).
Результаты
Метод «изоляции». Анализ экспериментальных данных показал близкие величины скорости разложения (по k, год-1) образцов основных фракций растительного материала в двух типах леса (таблица 2). Исключение составляет молодняк зеленомошного сосняка: более высокие показатели константы разложения опада и подстилки обеспечивает значительное участие листьев берёзы и осины в составе опадающей фитомассы.
Отсутствие существенных различий по скорости разложения органического вещества (ОВ) в различных типах леса объясняется близким биохимическим составом растительных остатков, в т.ч. высоким содержанием фенолов, лигнина (28- 34%), целлюлозы (34 -59%), при низкой концентрации азота, фосфора, кальция (Кулагина, 1978; Гирс, 1998 и др.).
Полученные оценки скорости разложения укладываются в пределы значений k, приводимых в литературе для сосновых насаждений схожих условий произрастания: древесина валежа – 0,015 – 0,044 год-1 и пней – 0,002 – 0,037 год-1, растительный материал подстилки и корневого детрита, соответственно, 0,089 и 0,072 год-1 (Krankina et al. 2002, Тарасов, 2002; Ведрова, 2005 и др.).
Насаждения, в которых проводились эксперименты, характеризуются близким запасом фитодетрита (рисунок 1) (Трефилова, 2006). Значительная доля (56-63%) ОВ растительных остатков в насаждениях, кроме перестойного сосняка зеленомошного, депонируется в мёртвой древесине валежа, сучьев, пней и сухостоя. Относительная роль подстилки с возрастом древостоя увеличивается от 18 и 17% в зеленомошном и лишайниковом молодняках, соответственно, до 44 и 25% в перестойных насаждениях, запас корневой мортмассы снижается – от 25 и 20% до 13 и 12%.
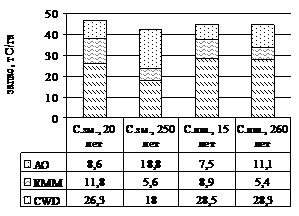
Рисунок 1 - Запас и структура фитодетрита изучаемых насаждений
«Потери» углерода при разложении фитодетрита, рассчитанные с учётом компонентного состава, в зеленомошных сосняках в среднем составляют 2,7 ц С га-1, что на 30% больше, чем в лишайниковых (таблица 3).
Основная часть (87-95%) продуктов разложения минерализуется и в форме СО2 возвращается в атмосферу. Роль отдельных компонентов фитодетрита в формировании минерализационного потока неравнозначна и изменяется с возрастом.
Таблица 2 - Скорость разложения растительных остатков (k, год-1)
Компоненты
|
С. зм.,
20 лет |
С. зм.,
250 лет |
С. лш.,
15 лет |
С. лш.,
260 лет |
|
|
|
|
|
Древесный опад |
0,291 |
0,154 |
0,201 |
0,113 |
Подстилка |
0,126 |
0,109 |
0,082 |
0,096 |
Опад+подстилка |
0,147 |
0,112 |
0,090 |
0,097 |
Опад корней |
0,08 |
0,08 |
0,08 |
0,08 |
Корневой детрит |
0,084 |
0,061 |
0,102 |
0,088 |
Отпад |
0,014 |
0,014 |
0,014 |
0,012 |
Грубые древесные остатки: |
|
|
|
|
в т.ч.: ветви |
0,014 |
0,014 |
0,014 |
0,014 |
Валёж |
0,013 |
0,013 |
0,008 |
0,008 |
Пни |
0,004 |
0,004 |
0,008 |
0,008 |
Сухостой |
- |
0,012 |
- |
0,012 |
Таблица 3 - Интенсивность минерализационного потока углерода в атмосферу, ц С га-1год-1
Интенсивность процессов, ц С га-1год-1 |
С. зм |
С. лш. |
20 лет |
250 лет |
15 лет |
260 лет |
Освободилось при разложении |
|
|
|
|
Опада и подстилка |
14,0 |
21,8 |
7,1 |
11,6 |
В т.ч. минерализация |
11,9 |
20,9 |
5,9 |
9,4 |
гумификация |
2,1 |
0,9 |
1,2 |
2,2 |
опада тонких корней и корневого детрита |
9,7 |
3,5 |
8,7 |
4,6 |
В т.ч. минерализация |
9,3 |
3,1 |
8,4 |
4,4 |
гумификация |
0,4 |
0,4 |
0,3 |
0,2 |
отпада текущего и прошлых лет |
2,8 |
1,9 |
2,9 |
2,5 |
Суммарные "потери" при разложении фитодетрита |
26,5 |
27,2 |
18,7 |
18,7 |
В т.ч. минерализация |
24,0 |
25,9 |
17,2 |
16,3 |
гумификация |
2,5 |
1,3 |
1,5 |
2,4 |
в т.ч. аккумуляция нового гумуса |
1,2 |
0,6 |
0,7 |
1,1 |
минерализация нового гумуса |
1,3 |
0,7 |
0,7 |
1,2 |
Минерализация С ПОВ гумуса |
2,6 |
0,6 |
1,8 |
1,1 |
Общий минерализационный поток |
26,7 |
26,6 |
19,0 |
17,5 |
При близкой массе фитодетрита молодняки разных типов леса различаются по интенсивности и удельной скорости гумификации. В зеленомошном сосняке удельная скорость гумификации составляет
5,4 мг С гС-1 год-1 и 2,5 ц С га-1 год-1 переходит в состав нового гумуса. В сосняке лишайниковом эти параметры составляют соответственно 3,3 мг С гС-1 год-1 и 1,5 ц С га-1 год-1.
В почвах спелых и перестойных древостоев со сформировавшимся составом гумуса современные процессы почвообразования идут по пути обновления периферической части гумусовых молекул (Фокин, 1974; Ганжара, 1979; Багаутдинов, Куватов, 1993 и др.). Аккумуляция новообразованного гумуса компенсирует минерализовавшийся подвижный С «старого» гумуса, в результате чего не происходит его дополнительного накопления в почве. В соответствии с этим ежегодная минерализация гумуса в почвах перестойных сосняков зеленомошного и лишайникового типов леса составляет соответственно 0,6 и 1,1 ц С га-1год-1(таблица3), за счет обновления «сработанного» гумуса новообразованными гумусовыми веществами запасы гумуса в почве не изменяются.
В молодых экосистемах (I – II класса возраста), формирующихся на месте вырубленного или (и) выгоревшего древостоя, регистрируется снижение запасов гумуса в результате интенсивной минерализации фитодетрита и подвижных форм собственно гумуса почв (Ведрова, 2005). Это подтверждается результатами наших исследований. В результате минерализации собственно гумуса почвы в атмосферу из молодняков поступает 2,6 и 1,1 ц С га-1 год-1. Около 50% потери гумуса компенсируется вновь синтезированным гумусом, «сработка» запаса гумуса в слое почвы 0-20 см составляет 1,4 и 1,1 ц С га-1 год-1, соответственно в зеленомошном и лишайниковом типах леса.
Согласно расчётам, гетеротрофное дыхание изучаемых насаждений в пределах одной группы типов леса мало изменяется с возрастом (таблица 3). Минерализационные «потери» ОВ почвы в зеленомошных сосняках на 30-34% выше, чем в лишайниковых.
Полученные величины укладываются в пределы усреднённых значений эмиссии СО2 с поверхности среднетаёжных почв (Кудеяров, 2005; Кудеяров, Курганова, 2005) и согласуются с оценками почвенного «дыхания» в средневозрастном сосняке черничном (Кайбияйнен, Ялынская, Софронова, 1999).
Метод «интеграции компонентов». Измерения эмиссии СО2 проводили три раза за сезон (первые декады июня, августа и сентября). Анализ полученных данных не выявляет значительных колебаний как между повторностями вариантов (V= 5-24%), так и показателями интенсивности эмиссии СО2.
Различия между максимальными и минимальными значениями, полученными в течение сезона, не превышают 18-30%. Исключением является молодняк лишайниковых сосняков. Интенсивность эмиссии СО2 в августе и сентябре на 67% ниже значений, зафиксированных в июне. Возможно, это является результатом пересыхания подстилки и верхнего минерального слоя почвы, и как следствие затухания процессов минерализации.
Всего с поверхности почвы 20-ти и 250-ти летних сосняков зеленомошного типа леса ежегодно выделяется 35 и 32 ц С га-1год-1, соответственно, в лишайниковых – 22 и 27 ц С га-1год-1 (таблица 4).
В исследуемых нами насаждениях основная часть потока углекислоты в атмосферу формируется в результате минерализации растительных остатков и подвижных форм гумуса почвы, на долю дыхания корней приходится 16-31%. Близкий порядок значений относительной доли дыхания корней (11-19%) получен для 80-100 летнего сосново-лиственного леса на дерново-подзолистой почве в Подмосковье (Ларионова и др., 2003). В смешанном сосновом (Pinussylvestris) лесу Швеции интенсивность корневого «дыхания» составляет 16 - 44% общего потока СО2 с поверхности почвы (Widen, Majdi , 2000).
Литературные данные, характеризующие роль дыхания корней сосны (Pinuselliottii, P. taedaL., P. SylvestrisL.), как источника эмиссии СО2, весьма противоречивы: от 27 до 73% (Ewel et.al. 1987; Maier, Kress, 2000; Цельникер, 2005 и др.). Часто вклад корней растений в почвенное дыхание принимают за 30%. На основе детального обзора установлено, что в лесных экосистемах «дыхание» корней растений в среднем составляет 45,8% от общей величины «дыхания» (Hanson, 2000).
Поскольку при оценке роли грубых древесных остатков в формировании минерализационного потока применение абсорбционного метода сопряжено с методическими сложностями, интенсивность минерализации древесного детрита определена, как и в предыдущем эксперименте, по запасу мёртвой древесины и скорости её разложения (таблица 4).
Таблица 4 - Интенсивность выделения СО2 с поверхности почвы и гетеротрофное дыхание в сосновых насаждениях зеленомошного и лишайникового типах леса, ц С га-1год-1
Потоки |
С.зм.,
20 лет |
С зм.,
250 лет |
С.лш.,
15 лет |
С лш.,
260 лет |
Интенсивность выделения СО2 с поверхности почвы, |
34,6 |
32,1 |
22,2 |
27,3 |
в т.ч. дыхание корней |
6,8 |
9,9 |
6,9 |
4,3 |
Минерализация грубых древесных остатков |
2,8 |
1,9 |
2,9 |
2,5 |
Гетеротрофное дыхание |
30,6 |
24,1 |
18,2 |
25,5 |
Сравнение двух методов оценки интенсивности гетеротрофного дыхания (см. таблицы 3 и 4) показало согласованность полученных результатов. Различия могут быть обусловлены неполной адекватностью состава разлагающихся подстилки и корневого детрита: при подготовке образцов для метода «изоляции»учитывалась вариабельность фракционного состава этих компонентов фитодетрита в пределах каждой пробной площади. При использовании метода «интеграции компонентов» фракционный состав подстилки и корневого детрита соответствовал лишь площадке, выбранной для эксперимента. Это не опровергает возможности использования любого из методов для изучения гетеротрофного дыхания экосистемы. В силу практических сложностей прямых измерений СО2-потока в ходе разложения грубых древесных остатков в лесных насаждениях с высоким запасом валежа и крупных веток предпочтение остается за методом «изоляции».
Выводы
- В ходе экспериментальных исследований в насаждениях разного возраста и типа леса оба используемых метода показали удовлетворительную сходимость результатов. Метод «интеграции компонентов» предпочтительнее использовать для оценки интенсивности гетеротрофного дыхания в экосистемах с низким запасом (или при отсутствии) грубых древесных остатков.
- Согласно полученным оценкам, интенсивность разложения фитодетрита в пределах одной группы изучаемых типов леса характеризуется близкими величинами. В зеленомошных сосняках Rh, примерно, на 30% выше, чем в лишайниковых. При близких валовых запасах фитодетрита, схожих значениях константы разложения, интенсивность освобождения углерода при разложении зависит от доли грубых древесных остатков в составе фитодетритного пула экосистемы.
- Основная масса углерода в гетеротрофном потоке формируется при разложении подстилки и корневого детрита, относительное участие грубых древесных остатков составляет 14-15% в сосняках лишайниковой и 7-10% в сосняках зеленомошной группы леса.
Автор выражает глубокую благодарность д.б.н. Э.Ф. Ведровой за ценные комментарии при обсуждении материала статьи.
Библиографический список
- Багаутдинов, Ф.Я. Трансформация меченых по углероду растительных остатков в черноземах типичных и серых лесных почвах / Ф.Я. Багаутдинов, Ю.Г. Куватов // Почвоведение. - 1993. - №3. - С. 25 - 31.
- Благодатский, С.А. Вклад дыхания корней в эмиссию СО2 из почвы / С.А. Благодатский, А. А. Ларионова, И.В. Евдокимов // Дыхание почвы. - Пущино: ОНТИ НЦБИ, 1993. - С. 26 - 32.
- Ведрова, Э.Ф. Разложение органического вещества лесных подстилок / Э.Ф Ведрова // Почвоведение. - 1997. - №2. - С. 216 - 223.
- Ведрова, Э.Ф. Баланс углерода в естественных и нарушенных южнотаёжных лесах Северной Сибири / Э.Ф. Ведрова, Л.С. Шугалей, В.Д. Стаканов // География и природные ресурсы. – 2002. - №4. - С. 92 - 99.
- Ведрова, Э.Ф. Деструкционные процессы в углеродном цикле лесных экосистем Енисейского меридиана: автореф. дис... док. биол. наук: 03.00.16. / Э.Ф Ведрова. - Красноярск, 2005. - 60 с.
- Ганжара, Н.Ф. О коэффициенте гумификации и методическом подходе к определению гумусного баланса в почвах / Н.Ф Ганжара // Почвоведение. - 1979. - №4. - С. 139 - 146.
- Гирс, Г.И. Содержание азота и зольных элементов в органах древесных растений России / Г.И. Гирс. – Красноярск,: 1998. - 76 с.
- Кайбияйнен, Л.К. Баланс углекислого газа в средневозрастном сосняке черничном / Л.К. Кайбияйнен, Е.Е. Ялынская, Г.И. Софронова // Экология. - 1999. - №4. - С. 271 - 275.
- Кауричев, И.С. Образование водорастворимых органических веществ в почвах как стадия превращения растительных остатков / И.С. Кауричев, И.М. Яшин // Известия ТСХА. - 1989. - Вып. 1. - С. 47 - 57.
- Куваев, В.Б. Естественное восстановление сосновых лесов среднего Енисея после рубок (опыт долговременного комплексного мониторинга) / В.Б. Куваев [и др.]. - М., 2001. - 314 с.
- Кудеяров, В.Н. Роль почвы в круговороте углерода / В.Н. Кудеяров // Почвоведение. - 2005. - №8. - С. 915 – 923.
- Кудеяров, В.Н., Дыхание почв России: анализ базы данных, многолетний мониторинг, общие оценки / В.Н. Кудеяров, И.Н. Курганова // Почвоведение. - 2005. - №9. - С. 1112-1121.
- Курганова, И.Н. Оценка потоков диоксида углерода из почв таёжной зоны России / И.Н. Курганова, В.Н. Кудеяров // Почвоведение. - 1998. - №9. - С. 1058 -1070.
- Кулагина, М.А. Биологическая продуктивность сосняков и круговорот макроэлементов / М.А. Кулагина // Продуктивность сосновых лесов. – М.: Наука, 1978. - С. 90 - 178.
- Ларионова, А.А. Дыхание корней и его вклад в эмиссию СО2 из почвы / А.А Ларионова [и др.].// Почвоведение. - 2003. - №2. - С. 183 - 194.
- Ларионова, А.А., Методы определения эмиссии СО2 из почвы / А.А Ларионова, Л.А. Иванникова, Т.С. Дёмкина // Дыхание почв. – Пущино: ОНТИ НЦБИ РАН, 1993. - С. 11-25.
- Ларионова, А.А. Динамика газообмена в профиле серой лесной почвы / А.А. Ларионова, Л.Н. Розонова, Т.И. Самойлов // Почвоведение. - 1988. - №11. - С. 68 – 74.
- Макаров, Б.Н. Изменения дыхания почвы и содержания углекислоты в приземном слое воздуха в течение суток / Б.Н. Макаров // ДАН СССР. - 1958. - Т.118. - №2. - С. 389 – 391.
- Макаров, Б.Н. Методы определения состава почвенного воздуха, интенсивности дыхания почвы и газообразных потерь азота почвы и удобрений / Б.Н. Макаров // Агрохимические методы исследования почв. - М.: Наука, 1975. - С. 331 - 344.
- Макаров, Б.Н. Газовый режим почв / Б.Н. Макаров. - М.: Агропромиздат, 1988. 104 с.
- Мина, В.Н. Опыт сравнительной оценки методов определения дыхания почв / В.Н. Мина // Почвоведение. - 1962. - №10. - С. 96 – 100.
- Наумов, А.В. Сезонная динамика и интенсивность выделения СО2 в почвах Сибири /А.В. Наумов // Почвоведение. - 1994. - №12. - С. 77 - 83.
- Пономарева, В.В. Методические указания по определению содержания и состава гумуса в почвах (минеральных и торфяных) / В.В. Пономарева, Т.А. Плотникова. – Л.: Наука, 1975. - 105с.
- Тарасов, М.Е. Методические подходы к определению скорости разложения древесного детрита / М.Е. Тарасов // Лесоведение. - 2002. - №5. - С. 32 - 38.
- Титлянова, А.А. Биологический круговорот углерода в травяных биогеоценозах / А.А. Титлянова. - Новосибирск: Наука, 1977. - 224 с.
- Трефилова, О.В. Органическое вещество почв сосняков Приенисейской Сибири / О.В. Трефилова // Вестник КрасГАУ. - 2006. - Вып.12. - С. 91 – 101.
- Фокин, А.Д. Включение органических веществ и продуктов их разложения в гумусовые вещества почвы / А.Д. Фокин // Известия ТСХА. – 1974. - Вып.6. - С. 99 - 110.
- Фокин А.Д. Участие различных соединений растительных остатков в формировании и обновлении гумусовых веществ почвы //Проблемы почвоведения. – М.: Наука, 1978. С. 60 - 64.
- Цельникер, Ю.Л. Дыхание корней и его роль в углеродном балансе древостоя / Ю.Л. Цельникер // Лесоведение. - 2005. - №6. - С. 11 - 18.
- Чагина, Е.Г. О балансе углерода при разложении опада в кедровниках Западного Саяна / Е.Г. Чагина // Вопросы лесоведения. - Красноярск, 1970. - Т.1. - С. 246 - 252.
- Чагина, Е.Г. Биологический круговорот в сосняках различной продуктивности / Е.Г. Чагина // Почвенные факторы продуктивности сосняков / под ред. В.В. Орловского. – Новосибирск: Наука, 1976. - С. 168 - 190.
- Шарков, И.Н. Определение интенсивности продуцирования СО2 почвой абсорбционным методом / И.Н. Шарков // Почвоведение. - 1984. - №7. - С. 136 - 143.
- Шарков, И.Н. Совершенствование абсорбционного метода определения выделения СО2 из почвы в полевых условиях / И.Н. Шарков // Почвоведение. 1987. - №1. - С. 127 - 133.
- Шарков, И.Н. Сравнительная характеристика двух модификаций адсорбционного метода определениядыхания почвы / И.Н. Шарков // Почвоведение. - 1987. - №10. - С. 153 - 157.
- Freijer, J.I. A comparison of field methods for measurement soil carbon dioxide evolution experiment and simulation / J.I. Freijer // Plant and Soil. - 1991. - V.135. - №1. - P. 133-142.
- Hanson, P.J. Separating root and soil microbial contributions to soil respiration: A review of methods and observations / P.J Hanson et al // Biogeochemistry. - 2000. - №48. - P. 115 – 146.
- Jenkinson, D.S. The turnover of soil organic matter in some of the Rothamsted classical experiments / D.S. Jenkinson, J.H. Rayner // Soil Science. - 1977. - V.123. - № 5. - Р. 298 - 305.
- Krankina, O.N. Coast woody debris in forest regions of Russia / O.N. Krankina et al // Can. J. For. Res. - 2002. - № 32. - Р. 768 - 778.
- Shvidenko, A. A synthesis of the impact of Russian forests on the global carbon budget for 1961 – 1998 / A. Shvidenko, S. Nilsson // Tellus. - 2003. - №55 - Р. 391 – 415.
- Vedrova, E.F. Net ecosystem production of boreal larch ecosystems on the Yenisei transect / E.F. Vedrova, F.I. Pleshikov, V.Y. Kaplunov // Mitigation and Adaptation Strategies for Global Change. - 2006. - V.11. - P. 173 – 190.
- Widen, B. Soil CO2 efflux and root respiration at three sites in a mixed pine and spruce forest: seasonal and diurnal variation / B. Widen, H. Majdi // Can J. For Res. - 2001. - V.31. - №5. - P. 786 – 796.
|


