|
|
|
Rus/Eng |
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
"Хвойные бореальной зоны" 2007г.,№4-5, с. 427-430 Особенности роста и формирования фитомассы древостоев ели в высокогорьях Южного Урала (на примере г. Малый Иремель) Нагимов З.Я. 1 , Бабенко Т.С. 1 , Шевченко И.Г. 1 , Рахманов И.В. 1 , Моисеев П.А. 2 1 Уральский государственный лесотехнический университет, Екатеринбург, Россия На разных высотных уровнях лесотундрового экотона в диапозоне от 1210 до 1360 м над уровнем моря рассмотрены зависимости высот и фракций фитомассы надземной и подземной частей деревьев в абсолютно сухом состоянии от их диаметров, особенности распределения количества деревьев по относительной высоте, определены запасы фитомассы древостоев. Выявлены различия в росте и структуре древостоев, связанные с высотой над уровнем моря. Средний диаметр и средняя высота древостоев закономерно уменьшаются с повышением высоты над уровнем моря. Коэффициент варьирования высот колеблется в пределах 17,2–38,2%, а диаметров – 26,3–66,8%. Средний возраст деревьев существенно уменьшается с увеличением высоты над уровнем моря от 36 лет наверху профиля до 95 лет внизу профиля. Наблюдается закономерное уменьшение полноты и густоты древостоев с повышением высоты над уровнем моря. На верхних уровнях связь высоты и диаметра деревьев ели на всем протяжении роста выражается прямой линией, а на нижних уровнях эта связь криволинейна. Это можно объяснить тем, что на верхних уровнях древостои сильно разрежены и по мере снижения высоты над уровнем моря увеличивается густота и полнота насаждений и, как следствие, конкурентные взаимоотношения между деревьями. По мере поднятия в гору меняется характер изменения высоты по ступеням диаметра и разряд высот. Максимальное количество деревьев на всех уровнях имеют относительную высоту от 41 до 60. В исследуемых насаждениях встречаются многоствольники, количество которых увеличивается с повышением высоты над уровнем моря. Общая фитомасса древостоев в абсолютно сухом состоянии по мере поднятия в гору уменьшается с 182,3 т/га на нижнем высотном уровне до 3,4 т/га – на верхнем. На исследуемом профиле уменьшение этого показателя на каждый метр высоты над уровнем моря составляет 1,19 т/га. В общей фитомассе древостоев доля корневых систем составляет 20,6–30,0%. There were described associations of trees heights and fractions of above and underground biomass in air-dry condition with their diameters, distribution of peculiarities of trees quantity upon their relative height on different altitudinal levels within 1210 and 1360 m a. s. l. There were determined actual reserves of the tree stands. The differences in the growth and structure of the stands linked with altitude were detected. The average diameter and average height of the stands regularly decrease with altitude increasing. The coefficient of variation of the heights varies within 17,2-38,2%, but diameter - 26,3-66,8%. The average tree age greatly decreases with altitude increasing from 36 years on the profile top upon 95 years at the bottom. Exists the natural reduction of the forest density and crowding level of the stands with altitude increasing. On top-levels relationship of spruce tree height and diameter during all growing is expressed with direct line, but on lower levels this relationship curvilinear. This possibly explains that on top-levels stands are strongly rarefied and as the altitude decreases the forest density and crowding level of the stands are increase and, as effect, competitive relations between trees are increase too. With the lifting uphill the nature of the height change on step of the diameter and category of the heights are change. The maximum amount of the trees on all levels have a relative height from 41 before 60. There are found multi-stem trees in the investigation area, the amount of which increases with altitude increasing. The total phytomass of the tree stands decreases with the lifting uphill from 182,3 tons/ha at the bottom levels to 3,4 tons/ha - on the top levels. In the investigation area reduction of this factor on each meter of the altitude is 1,19 tons/ha. In the total stands biomass share of the root systems is 20,6-30,0%. Теоретическая часть В связи с существенным потеплением климата нашей планеты в 20 веке, значительно увеличился интерес мировой научной общественности к изучению реакции экосистем на это явление. Наиболее чувствительными к изменению климатической обстановки являются экосистемы, расположенные в высокоширотных и высокогорных районах. Высокогорные экосистемы в настоящее время остаются наименее изученными в лесоводственно-таксационном отношении. В то же время в рассматриваемом аспекте вопросы формирования насаждений, роста и дифференциации деревьев в условиях верхнего предела произрастания леса, приобретают особую актуальность. Исследования их важны для оценки динамики верхней границы леса и биосферной роли насаждений, произрастающих в этих условиях. В частности, повышение верхней границы леса в горах свидетельствует о расширении площадей насаждений, обеспечивающих длительное консервирование углерода. Для оценки углерододепонирующей роли этих насаждений необходимы целенаправленные исследования их роста и продуктивности. Объекты и методика исследований Исследования проводились по методике международного проекта INTAS-01-0052. На юго-западном склоне горы Малый Иремель был заложен высотный профиль. Для его закладки подбирался участок склона с хорошо сформированными почвами и типичной для данного района растительностью. На профиле фиксировались четыре высотных уровня: первый (верхняя часть границы леса) – на высоте 1360 м над уровнем моря, второй (средняя часть границы леса) – на высоте 1300 м над уровнем моря, третий (нижняя часть границы леса) – на высоте 1260 м и четвертый (верхняя часть горных лесов) – на высоте 1210 м. На этих уровнях согласно методике вышеуказанного проекта закладывались по 5 - 6 макроплощадок размером 20?20 м. В пределах высотного уровня макроплощадки располагались таким образом, чтобы их центры отстояли друг от друга на расстоянии не более 5-10 м в вертикальном направлении и не более 50-100 м в горизонтальном. На всех макроплощадках для каждого дерева устанавливались: номер, порода, точное местоположение, форма ствола (одноствольная, многоствольная), высота, диаметр у основания и на высоте груди, диаметр и протяженность кроны, а также возраст по буровым кернам. Результаты и обсуждение Основные таксационные показатели исследованных древостоев представлены в таблице 1. Таблица 1 - Таксационная характеристика древостоев по высотным уровням
Наблюдается закономерное уменьшение полноты и густоты древостоев с повышением высоты над уровнем моря. Эта характерная особенность верхней границы леса описана во многих работах отечественных и зарубежных исследователей.
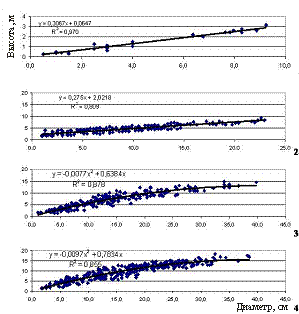
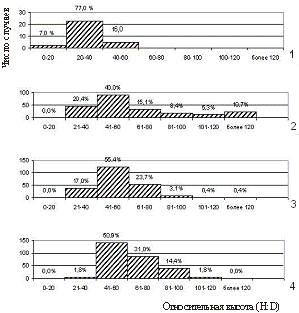
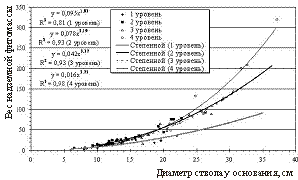
снижения высоты над уровнем моря. Об этом свидетельствуют и расположение экспериментальных точек на графиках и значения коэффициентов при х2 в уравнениях зависимости. При снижении высоты над уровнем моря увеличивается густота и полнота насаждений, а следовательно, и конкурентные взаимоотношения между деревьями, которые являются основной причиной нарушения постоянства соотношений между диаметром и высотой деревьев. При одинаковом диаметре высота деревьев закономерно снижается с повышением высоты над уровнем моря. Таким образом, по мере поднятия в гору меняется характер изменения высоты по ступеням диаметра и разряд высот. Таким образом, наличие в исследуемых насаждениях достаточно обособленных биогрупп, многоствольных форм и отдельно растущих деревьев обуславливает достаточно широкий диапазон варьирования относительных высот и неравномерность распределения деревьев по ступеням данного показателя. Средние значения относительной высоты деревьев, рассчитанные по десятилетиям возраста, свидетельствуют, что степень напряженности конкурентных взаимоотношений между деревьями закономерно снижается с увеличением высоты над уровнем моря. Рисунок 3 – Зависимость надземной фитомассы деревьев от их диаметра Таблица 2 – Запасы фитомасс древостоев по фракциям на различных высотных уровнях
Примечание: Фитомасса древостоев: в числителе – общая (ели и березы), в знаменателе – ели В общей фитомассе древостоев доля корневых систем составляет 20,6–30,0 %. Наблюдаются определенные зависимости в изменении соотношенийфракций фитомассы древостоев, связанные с высотой их произрастания относительно уровня моря: по мере продвижения в гору уменьшается доля хвои и сухостоя и повышается доля корней. Заключение В целом результаты исследований свидетельствуют, что на верхней границе леса даже при незначительных перепадах высот над уровнем моря существенно меняются характер распределения деревьев по таксационным признакам и формы взаимосвязей между диаметром и высотой деревьев. Для еловых древостоев характерна значительная амплитуда изменения таксационных показателей и общей фитомассы. В зависимости от высоты над уровнем моря наблюдаются закономерное изменение как абсолютных значений фракций фито- массы, так и соотношений их между собой. БИБЛИОГРАФИЧЕСКИЙ СПИСОК
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||