|
"Хвойные бореальной зоны" 2007г.,№2-3, с.
АЛЛОЗИМНАЯ ИЗМЕНЧИВОСТЬ МОЖЖЕВЕЛЬНИКА КАЗАЦКОГО JUNIPERUS SABINA L. НА ЮЖНОМ УРАЛЕ
Янбаев Ю.А., Редькина Н.Н., Муллагулов Р.Ю.
Сибайский институт Башкирского государственного университета, г. Сибай,
Республика Башкортостан, г. Сибай, ул. Белова, д. 21, т (34775) 3-43-93, yanbaev_ua@mail.ru
C использованием полиакриламидного диск-электрофореза и изоферментов в качестве генетических маркеров в местообитании можжевельника казацкого Juniperussabina L. изучена клональное разнообразие и генетическая изменчивость вида. Из-за широкой распространенности в Республике Башкортостан вид не отнесен к категории редких и исчезающих растений. Однако в настоящее время в условиях Башкирского Зауралья наблюдается сокращение площадей и ухудшение состояния можжевельников. Цель работы - разработка подходов для сохранения и рационального использования генетических ресурсов J. sabina на основе исследования его генотипической структуры. Для оценки клонального разнообразия и генетической изменчивости растения использованы 11 локусов (Aat-1, Dia-1, Gdh-1, Mdh-1, Mdh-2, Lap-1, Lap-2, 6Pgdh-1, Fdh-1, Skdh-1, Est-1). Из их числа полиморфными были 10 (за исключением Gdh-1), среднее число аллелей на локус составило 2.2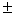 0.2, ожидаемая и наблюдаемая гетерозиготность составили значения Не = 0.327 0.2, ожидаемая и наблюдаемая гетерозиготность составили значения Не = 0.327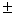 0.054 и Но =0.323 0.054 и Но =0.323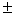 0.055, соответственно. Можжевельник казацкий обладает относительно высоким генетическим разнообразием, по сравнению с другими видами с близкими биоэкологическими свойствами. Стратегия сохранения генофонда in situ должна включать охрану местообитаний с наибольшим клональным и генетическим разнообразием. Выявленная генетическая структура в местообитании можжевельника казацкого показывает, что наибольший интерес представляет использование большого числа куртин небольшого размера, каждая из которых обладает отдельным генотипом. Уникальная древовидная форма, произрастающая в большой куртине с кустовидными и стланиковым можжевельниками, образует вместе с ними один гигантский клон с одинаковым генотипом. 0.055, соответственно. Можжевельник казацкий обладает относительно высоким генетическим разнообразием, по сравнению с другими видами с близкими биоэкологическими свойствами. Стратегия сохранения генофонда in situ должна включать охрану местообитаний с наибольшим клональным и генетическим разнообразием. Выявленная генетическая структура в местообитании можжевельника казацкого показывает, что наибольший интерес представляет использование большого числа куртин небольшого размера, каждая из которых обладает отдельным генотипом. Уникальная древовидная форма, произрастающая в большой куртине с кустовидными и стланиковым можжевельниками, образует вместе с ними один гигантский клон с одинаковым генотипом.
Clonal diversity and genetic variation within a habitat of Juniperus sabina L. were investigated using polyacrylamide disc-electrophoresis and isozymes as genetic markers. Because of its vast geographic range, the plant is not considered threatened in the Republic of Bashkortostan. However, within Bashkirian trans-Urals there has been a substantial decline in both the distribution of the juniper and the сonditions of its colonies. The purpose of the investigation was to propose a conservation strategy based on the observed genotypic structure. Eleven allozyme loci (Aat-1, Dia-1, Gdh-1, Mdh-1, Mdh-2, Lap-1, Lap-2, 6Pgdh-1, Fdh-1, Skdh-1, Est-1) were used to examine genetic and clonal diversity in a natural population of the juniper. Ten loci (90.9 %), excluding Gdh-1, were polymorphic, with a mean number of alleles per locus of 2.2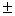 0.2. Mean expected heterozygosity within the species (He) was 0.327 0.2. Mean expected heterozygosity within the species (He) was 0.327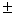 0.054, and observed heterozygosity (Ho) was 0.323 0.054, and observed heterozygosity (Ho) was 0.323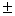 0.055. Compared to species having similar life-history traits, J. sabina displayed high genetic diversity. The most suitable strategy for conservation of genetic diversity in situ of the species is the protection of habitats with highest clonal diversity and genetic variation. The observed patterns of genotypic structure indicates that management for conservation of genetic variability should not only aim to preserve large colonies but also as many of the small bushes (with unique genotypes) as possible. An unique large group of plants of columnar growth form, mat-forming and shrub-like individuals represents one clone with the same genotype. 0.055. Compared to species having similar life-history traits, J. sabina displayed high genetic diversity. The most suitable strategy for conservation of genetic diversity in situ of the species is the protection of habitats with highest clonal diversity and genetic variation. The observed patterns of genotypic structure indicates that management for conservation of genetic variability should not only aim to preserve large colonies but also as many of the small bushes (with unique genotypes) as possible. An unique large group of plants of columnar growth form, mat-forming and shrub-like individuals represents one clone with the same genotype.
Введение
Сохранение генетического разнообразия популяций в последние десятилетия стало одной из глобальных проблем человечества, так как именно оно определяет потенциал для адаптации и эволюции. Древесные растения выделяются спецификой структуры внутривидовой изменчивости [1], что необходимо учитывать при разработке мер сохранения генофонда природных популяций методами ex situ и in situ [2]. У каждого вида в течение многих поколений сложилась определенная структура внутривидовой изменчивости, игнорирование чего может привести к множеству отрицательных последствий [3]. В настоящее время во всем мире и России, благодаря широкому использованию изоферментных маркеров, исследована генетическая изменчивость большей части видов древесных растений [4, 5, 6]. К сожалению, аналогичных данных, в том числе с использованием аллозимов в качестве генетических маркеров, для видов можжевельника практически нет. Лишь в последние годы появляются публикации (все они за пределами России) по изучению можжевельников с применением маркеров ядерной и хлоропластной ДНК [7, 8, 9, 10].
В Башкортостане из трех видов можжевельника особенно распространенным являетсяJuniperussabinaL. Он является одним из наиболее важных компонентов чрезвычайно уязвимых горно-степных экосистем Башкирского Зауралья. Несмотря на то, что он включался во 2-е издание Красной книги Республики Башкортостан, из-за неприхотливости и большой устойчивости к антропогенному воздействию является относительно распространенным в регионе. Вид имеет огромное значение для сохранения целостности этих экосистем. Растения, разрастаясь огромными куртинами, препятствуют эрозии горных почв и восстанавливают их после выпаса скота. Можжевельник казацкий относится к реликтовым элементам флоры доледниковой эпохи на Южном Урале. Он образует множество морфологических форм и разновидностей, имеющих большие перспективы для ландшафтного дизайна. Кроме того, в Башкирском Зауралье в настоящее время ведутся работы по созданию особо охраняемых природных территорий, однако они до сих пор остаются без необходимого научного обоснования в отношении сохранения генофонда J. sabina. Эти причины обуславливают актуальность проведенного нами генетического анализа внутривидовой изменчивости можжевельника казацкого.
Цель проведенного исследования – электрофоретическое изучение аллозимной изменчивости стланиковой и древовидной форм можжевельника казацкого на территории Башкирского Зауралья.
Экспериментальная часть
Район исследования находится на юго-востоке Республики Башкортостан на территории Хайбуллинского района и представляет сильно расчлененный горно-лесостепной ландшафт на восточном макросклоне южноуральских гор. На южном склоне горы в долине р. Зилаир нами было выбрано местообитание можжевельника казацкого и на территории 1 га была проведена инвентаризация куртин растения с определением их общей высоты, площади и конфигурации. В начале мая 2007 г. были собраны ветки для лабораторных анализов.
Разделение изоферментов проведено методом диск-электрофореза в вертикальных пластинах 7.5 %-го полиакриламидного геля с рН разделяющего геля 8.9 [11, 12] с использованием трис-глицинового электродного буфера с рН 8.3. Изучена изменчивость аллозимов аспартатаминотрансферазы (AAT, E.C. 2.6.1.1.), диафоразы (DIA, 1.6.4.3), глутаматдегидрогеназы (GDH, 1.4.1.2), малатдегидрогеназы (MDH, 1.1.1.37), лейцинаминопептидазы (LAP, 3.4.11.1), 6-фосфоглюконатдегидрогеназы (6-PGD, 1.1.1.44), формиатдегидрогеназы (FDH, 1.4.1.2), шикиматдегидрогеназы (SKDH, 1.1.1.25) и неспецифических эстераз (EST, 3.1.1…).
Для экстракции ферментов использован 0.1 М трис-HCl (рН 8.0) буфер, содержащий 17 % сахарозы, 0.1 % 2-меркаптоэтанола, 0.05 % диэтилдитиокарбомата натрия. Навеску кусочков стеблей в 200 мг растирали в фарфоровой ступке с добавлением 1 мл охлажденного экстрагирующего буфера. Для связывания полифенолов перед этим добавляли около 200 мг нерастворимого поливинилпирролидона Polyclar AT. Гомогенат центрифугировали в течение 20 минут при 15-17 тыс. об/мин в холодильнике. Гистохимическое выявление зон ферментативной активности в гелях осуществлено по стандартным методикам [13]. Для обозначения ферментов и локусов применена одна из общепринятых номенклатур [14]. Аллели обозначались арабскими цифрами, возрастающими по мере уменьшения электрофоретической подвижности их изоферментов. Гипотезы о генетическом контроле ферментов были выдвинуты на основе анализа фенотипов изоферментов (пространственной разобщенности зон активности в гелях, образования гибридных изоферментов у гетерозигот), статистического тестирования различий выявленного соотношения фенотипов и их теоретически ожидаемых частот, а также опубликованных данных по механизмам наследования изоферментов у других видов можжевельника [7].
Многолокусные генотипы определялись с использованием программы GSЕD [15]. При анализе уровня изменчивости выборок использованы стандартные показатели, определяемые компьютерной программой BIOSYS-1 [16]: частоту аллелей и их среднее число на локус, долю полиморфных локусов, наблюдаемую и ожидаемую гетерозиготность.
Результаты и обсуждение
Специфика местообитаний можжевельника в Башкирском Зауралье заключается прежде всего во фрагментарности его распространения. Куртины располагаются среди горных степей, лугов и скалистых обнажений в целом мозаично. Границы зарослей определяются, видимо, микрогетерогенностью среды (температурный и ветровой режимы, увлажненность и интенсивность освещения, развитость почвенного покрова и т.д.). На 1 га детально исследованной территории выявлено 32 куртины со средней площадью 9.82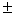 1.92 м2. (с изменениями в пределах 0.6-26.6 м2., коэффициент вариации 78.3%) и общей площадью 137.7 м2./га. Доминируют (91.35%) округлые по форме (с соотношением сторон менее 2 : 1) и вытянутые куртины, а их распределение по площади близко к гиперболической кривой - преобладают участки небольшого размера. Лишь в глубоких логах вне пробной площади выявлены сплошные заросли площадью в несколько гектаров. 1.92 м2. (с изменениями в пределах 0.6-26.6 м2., коэффициент вариации 78.3%) и общей площадью 137.7 м2./га. Доминируют (91.35%) округлые по форме (с соотношением сторон менее 2 : 1) и вытянутые куртины, а их распределение по площади близко к гиперболической кривой - преобладают участки небольшого размера. Лишь в глубоких логах вне пробной площади выявлены сплошные заросли площадью в несколько гектаров.
Установлено, что практически каждая небольшая куртина на пробной площади электрофоретически гомогенна по спектру изоферментов, т.е. представляет отдельный генотип. Видимо, распространение можжевельника происходит не только за счет увеличения размеров куртин из-за вегетативного возобновления, но и в значительной степени семенным способом. Дополнительным свидетельством этому заключению может быть соответствие наблюдаемых распределений их теоретически ожидаемым частотам. Несмотря на относительно малую численность выборки куртин (исследованы 11 генотипов), ни по одному из использованных 10 изменчивых локусов не отмечено нарушение правила Харди-Вайнберга. Частоты аллелей и гетерозиготность выборки клонов стланиковой формы приведены в таблице 1.
Таблица - Частота аллелей и гетерозиготность J. sabinaL.
Локусы |
Частота аллелей |
Гетерозиготность |
1 |
2 |
3 |
ожидаемая |
наблюдаемая |
Dia-1 |
0.778 |
0.222 |
- |
0.346 |
0.444 |
Mdh-2 |
0.833 |
0.167 |
- |
0.278 |
0.333 |
Mdh-1 |
0.111 |
0.889 |
- |
0.198 |
0.222 |
Aat-1 |
0.056 |
0.944 |
- |
0.105 |
0.111 |
Fdh-1 |
0.333 |
0.667 |
- |
0.444 |
0.444 |
Gdh-1 |
1.000 |
- |
- |
0 |
0 |
Skdh-1 |
0.500 |
0.500 |
- |
0.500 |
0.556 |
Lap-2 |
0.111 |
0.778 |
0.111 |
0.370 |
0.222 |
Lap-1 |
0.056 |
0.611 |
0.333 |
0.512 |
0.556 |
Est-1 |
0.333 |
0.667 |
- |
0.444 |
0.444 |
6Pgdh-1 |
0.056 |
0.889 |
0.056 |
0.204 |
0.222 |
Из 11 исследованных локусов лишь Gdh-1 был мономорфным. Слабый полиморфизм выявлен для Aat-1, в остальных локусах обнаружены по несколько аллелей, два из которых имеют сопоставимые частоты. По этой причине получены относительно высокие показатели генетического разнообразия – доля полиморфных локусов Р = 90.9, среднее число аллелей на локус А = 2.2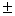 0.2, ожидаемая гетерозиготность Не = 0.327 0.2, ожидаемая гетерозиготность Не = 0.327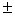 0.054, Но = 0.323 0.054, Но = 0.323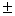 0.055. Полокусные значения гетерозиготности подтверждают этот вывод. В единственном другом исследовании можжевельников с использованием аллозимов [7] показано, что J. rigida Sieb. Et. Zuss из Кореи полиморфна по 16 из 22 изученных локусов (72.7 %), а каждая отдельная популяция была изменчива по 45.4 % генов, с изменениями от 36.4 до 54.6 %. В среднем на один локус выявлено 1.61 аллелей, наблюдаемая гетерозиготность составила значение 0.187. Не намного J. rigida уступал другой эндемичный вид J. coreana Nakai (Р = 54.6 %, А = 2.42, Не = 0.199). Таким образом, и в цитируемой работе подтверждается результат, полученный нами для J. sabinaL., о сравнительно высоком по сравнительно с другими древесными видами [17] уровне генетического разнообразия можжевельников. Он может быть обусловлен как эколого-географической широтой ареала вида, так и его биоэкологическими свойствами, т.ч. способностью «консервировать» генофонд благодаря вегетативному размножению. 0.055. Полокусные значения гетерозиготности подтверждают этот вывод. В единственном другом исследовании можжевельников с использованием аллозимов [7] показано, что J. rigida Sieb. Et. Zuss из Кореи полиморфна по 16 из 22 изученных локусов (72.7 %), а каждая отдельная популяция была изменчива по 45.4 % генов, с изменениями от 36.4 до 54.6 %. В среднем на один локус выявлено 1.61 аллелей, наблюдаемая гетерозиготность составила значение 0.187. Не намного J. rigida уступал другой эндемичный вид J. coreana Nakai (Р = 54.6 %, А = 2.42, Не = 0.199). Таким образом, и в цитируемой работе подтверждается результат, полученный нами для J. sabinaL., о сравнительно высоком по сравнительно с другими древесными видами [17] уровне генетического разнообразия можжевельников. Он может быть обусловлен как эколого-географической широтой ареала вида, так и его биоэкологическими свойствами, т.ч. способностью «консервировать» генофонд благодаря вегетативному размножению.
Особый интерес с точки зрения охраны и рационального использования генетических ресурсов можжевельника казацкого имеет обнаруженная ранее [18] древовидная форма. Если обычно растение имеет вид стелющегося кустарника с приподнятыми ветвями высотой до 0.7-1.0 м, древовидный можжевельник имеет высоту до 4 м высоты. Примечательно, что он выделяется голубовато-зеленым цветом хвои, что дает дополнительные аргументы для обоснования перспективности в ландшафтном озеленении. В пределах имеющейся в районе исследований одной большой куртины нами выявлено 7 экземпляров древовидного можжевельника, взятые для электрофоретического анализа, с относительно большим (до 20 см) диаметром на высоте груди. Также здесь представлены особи относительно небольшого размера, но древовидность ствола которых уже не вызывает сомнения. Эти экземпляры находятся в центре плотных зарослей кустовидного и стелющегося можжевельника, среди которых из разных сторон куртины для анализа частоты встречаемости аллозимов были взяты 8 образцов.
Установлено, что все 7 образцов древовидного можжевельника имеют один и тот же уникальный «генетический портрет», т.е. являются представителем одного клона, гетерозиготного по локусам Dia-1 и Est-1. Таким же генотипом обладали стланиковые и кустовидные 7 исследованных экземпляров. Лишь восьмой образец был гетерозиготным по локусам Fdh-1, Skdh-1, Lap-1, Lap-2 и Est-1, и гомозиготным по Dia-1. Другими словами, куртина с древовидным можжевельником является гигантским клоном с небольшой примесью других генотипов.
Выводы
Полученные результаты доказывают перспективность генетических маркеров для сохранения и рационального использования генетических ресурсов можжевельника казацкого. Аллозимы оказались эффективным средством для идентификации клонов и генотипов можжевельника казацкого. Большая встречаемость и небольшой размер куртин можжевельника может свидетельствовать о том, что их возраст в исследованном районе сравнительно небольшой. Именно местообитания с наибольшим разнообразием (числом генотипов) представляют, видимо, первоочередной интерес при выборе объектов сохранения генофонда in situ, а также при отборе образцов для создания коллекции клонов ex situ. Полученные нами результаты помогут внести поправки для проведения работ по сохранению генотипа древовидного можжевельника вне этих местообитаний (методы ex situ, например, интродукция и реинтродукция в других местностях). Сравнительно дорогостоящие, требующие больших затрат времени и усилий работы по перемещению и размножению всех древовидных экземпляров приведут к тому, что сохранены и использованы будут образцы только одного и того же клона. Что также важно – нам удалось показать, что клонами являются и экземпляры можжевельника казацкого небольших размеров, у которых из-за относительно небольшого возраста еще не выявляется «древовидность» формы. Это расширяет возможности отбора образцов, а также позволит более успешно проводить работы по размножению клона ex situ.
Библиографический список
- Алтухов, Ю.П. Внутривидовое генетическое разнообразие: мониторинг и принципы сохранения / Ю.П. Алтухов // Генетика. - 1995. - Т. 31. - С. 1333-1357.
- Корочкин Л.И. Генетика изоферментов./ Л.И. Корочкин, [и др.]. - М.: Наука, 1977. - 275 с.
- Мамаев, С.А. О популяционном подходе в лесоводстве / С.А. Мамаев, Л.Ф. Семериков, А.К. Махнев // Лесоведение. – 1988. - № 1. – С. 3-9.
- Мамаев, С.А. Охрана генофонда древесных растений в России / С.А. Мамаев, А.К. Махнев, А.И. Ирошников // Материалы международного симпозиума “Лесная генетика: охрана, воспроизводство и рациональное использование генетических ресурсов” (г. Уфа, 4-11 августа 1991). - Уфа: 1994. – С. 24-36.
- Милютин, Л.И. Исследование популяционных генофондов лесообразующих видов древесных растений Сибири с целью сохранения, отбора и рационального использования ценных популяций / Л.И. Милютин // Динамика генофондов растений, животных и человека: отчетная конф. по программе фундаментальных исследований РАН. – М.: ИОГен РАН, 2005. – С. 87-89.
- Мулдашев, А. Древовидный можжевельник / А.Мулдашев, С.Кучеров // Табигат. - № 1 (36). – 2005. – С. 24-25.
- Политов, Д.В. Генетика популяций кедровых сосен/ Д.В.Политов, М.М Белоконь, Ю.С. Белоконь [и др.] // Генетика в России и в мире. – 2006. – С. 156.
- Davis, B.J. Disc electrophoresis. 11. Methods and application to human serum proteins / B.J. Davis // Ann. New York Acad. Sci. - 1964. - V. 121. P. 404-427.
- Gillet, E. GSED: Genetic Structures from Electrophoresis Data. Computer program and user's manual / E. Gillet // http://www.uni-forst.gwdg.de/forst/fg/software.htm, 1998.
- Goncharenko, G.G. Genetic resources of pine, spruce and fir species in the former Soviet Union: analysis of their genepools, phylogenetic relationships and genome organization / G.G. Goncharenko, , et al. // Sustainable forest genetic resources programmes in the Newly Independent States of the former USSR. Proceedings of a workshop, 23-26 September 1996, Belovezha, Belarus (Goncharenko G.G., Turok T., Gass T. and Paule L., eds). Copublished by Arbora Publishers, Zvolen, Slovakia and International Plant Genetic Resources Institute, Rome, Italy. – 1998. – P. 83-101.
- Hamrick, J.L. Factors influencing levels of genetic diversity in woody plant species / J.L.Hamrick, M.J.W. Godt, S.L. Sherman-Broylers // New Forest. - 1992. - № 6. - P. 95-124.
- Man, K.H. Genetic diversity and population structure of Juniperus rigida (Cupressaceae) and Juniperus coreana / K.H.Man, W.H. Hong // Evolutionary Ecology. – 2000. – V. 14. – P. 87-98.
- Meloni, M. Genetic variation in five Mediterranean populations of Juniperus phoenicea as revealed by inter-simply sequence repeat (ISSR) markers / M.Meloni, Et al. // Ann. Bot. – 2006. – V. 97. – P. 299-304.
- Merwe, M.V. Spatial and temporal aspects of the genetic structure of Juniperus communis populations / M.V.Merwe, et al. // Mol. Ecol. – 2000. – V. 9. P. 379-386.
- Ornstein, L. Disc-electrophoresis. I. Background and theory / L. Ornstein // Ann. New York. Acad. Sci. - 1964. V.121. - P. 321-349.
- Prakash, S. A molecular approach to the study of genetic heterozygosity in natural populations. IV. Patterns of genic variation in central, marginal and isolated populations of Drosophila pseudoobscura / S. Prakash, R.C. Lewontin, J.L. Hubby // Genetics. - 1969. - V.61. - P. 841-858.
- Swofford, D.L. BIOSYS-1: a FORTRAN program for the comprehensive analysis of electrophoresis data in population genetics and systematics / D.L.Swofford, R.B. Selander // J. Heredity. - 1981. - V.72. - P.281-283.
- Terry, R.G. Genetic variation in chloroplast and nuclear ribosomal DNA in Utah juniper (Juniperus osteosperma, Cupressaceae): evidence for interspecific gene flow / R.G.Terry, R.S. Nowak, R.J. Taush // Am. J. Bot. – 2000. – V. 87. – P. 250-258.
|


