|
"Хвойные бореальной зоны" 2007г.,№2-3, с.
ГИДРОХОРИЯ КАК ФАКТОР ГЕНЕТИЧЕСКОЙ ИНТЕГРАЦИИИ ДИФФЕРЕНЦИАЦИИ ПОПУЛЯЦИЙ ХВОЙНЫХ ДРЕВЕСНЫХ РАСТЕНИЙ
Санникова Н.С., Санников С.Н.
Ботанический сад Уральского отделения Российской Академии Наук, Екатеринбург, Россия
620144, г.Екатеоинбург, ул. 8 марта, 202,т. (343) 2103859, stanislav.sannikov@botgard.uran.ru
На примере лесообразующих видов хвойных Северной Евразии – сосны обыкновенной (PinussylvestrisL.) и ели европейской (Piceaabies (L.)Karst.) – рассмотрены результаты сравнительного экспериментального и натурного изучения скорости и дальности анемохорного и гидрохорного распространения семян и расселения популяций в равнинных и горных лесах. Установлено, что максимальная скорость расселения самосева сосны в центре ее ареала на оптимальном минерализованном субстрате заброшенных пашен не превышает 90-140 км/1000 лет, а скорость расселения самосева того же вида и ели европейской по течению горной реки Ломницы (Украинские Карпаты) достигает 1700-2000 км/1000 лет, т.е. на порядок выше. В подтверждение гипотезы (Санников, Санникова, 2007) о приоритете гидрохории семян в процессе расселения видов хвойных показаны высокая степень сходства аллозимной структуры популяций Pinussylvestris, расположенных на трансектах вдоль русел рек (генетические дистанции Неи (Nei, 1978) – от 0,004 до 0,008), и в 1,5-2,5 раза большие различия на трансектах, пересекающих их водоразделы, особенно в горных условиях. Сделан вывод о том, что гидрохория семян многих видов хвойных по течению рек – один из ведущих факторов, определяющих пути и скорость миграции, а также степень геногеографической интеграции и дифференциации их популяций в плейстоцене.
Results of the comparative experimental and large-scale studies concerned with the anemochorous and the hydrochorous migration of seeds and dispersal of populations in plain and montane forests have been considered by the example of forest-forming coniferous species – the common pine (Pinus sylvestris L.) and the Norway spruce (Picea abies (L.)Karst.) – in Northern Eurasia. It was found that the maximum dispersal rate of the pine selfsown at the center of the range is not over 90-140 km in 1000 years on the optimal mineralized substrate of abandoned agricultural lands, whereas the dispersal rate of the selfsown of the common pine and the Norway spruce with stream of the mountain river Lomnitsa (the Ukrainian Carpathians) is as large as 1700-2000 km/1000 years, i.e. one order of magnitude higher. The hypothesis (Sannikov, Sannikova 2007) for the priority of the seed hydrochory in the dispersal of coniferous species has been supported by the high similarity of the allozymic structure of Pinus sylvestris populations in transects along river beds (Nei (1978) genetic distances of 0.004 to 0.008) and largely different allozymic structures (1.5-2.5 times) in transects going across the river basins, especially in mountains. It is concluded that the hydrochory of seeds of many coniferous species along rivers is one of the major factors determining the paths and the rate of migration, as well as the genogeographic integration or differentiation of their populations in the Pleistocene.
Введение
Изучение способов, путей и темпов расселения популяций древесных растений в послеледниковые эпохи – одна из ключевых, но недостаточно разрабатываемых проблем современной эволюционной биологии.
К настоящему времени благодаря бурному прогрессу молекулярно-генетических методов довольно широко и разносторонне изучена географическая изменчивость состава изозимов и частью гаплотипов ДНК многих видов хвойных, в частности сосны обыкновенной Prus-Glowacki et. al., 1993; Krutovskii, Bergman, 1995; Гончаренко, 1999; Semerikov et. al., 2002; Потенко, 2004; Ларионова, 2004). В пределах ареалов видов выявлены географические особенности дифференциации аллозимной структуры, градиенты и границы популяций сосны обыкновенной , а также гаплотипов ДНК популяций лиственниц (Санников, Петрова, 2003; Семериков, 2007). Однако феноменологически установленные геногеографические закономерности структуры популяций не получили достаточной палеогеографической и экологической интерпретации. К числу основных факторов, изучение которых могло бы во многом способствовать пониманию закономерностей дивергенции популяций, выявлению центров их происхождения и филогенетических связей, следует отнести пути и темпы их миграции в различных частях ареала.
Скорость расселения популяций хвойных большинство авторов явно или неявно связывает со скоростью их анемохории (Frenzel, 1968; Удра, 1988; Lang, 1994). Однако весьма быстрое распространение видов хвойных в межлелниковые эпохи – со скоростью до 360-600 км/1000 лет, установленное палинологами (Birks, 1989; Lang, 1994; Kremeneckii et. al., 1998; Cheddadi et. al., 2006), не может быть объяснено только анемохорией их семян, так как ее средняя скорость не превышает 50-60 км/1000 лет (Шиманюк, 1955; Удра, 1988; Санников, 1992). Между тем ранее нами была выявлена высокая плавательная способность семян сосны (с крылатками) в речной воде (Санников, 1976).
На этом основании выдвинута гипотеза о возможности дальнего водного транспорта семян некоторых видов хвойных по течению рек (Санников, Санникова, 2007). Следствием этой гипотезы должны быть сходство генофонда смежных поселений сосны, расположенных вдоль русел рек, а с другой стороны, его сравнительно большая степень дифференциации в направлении поперек водоразделов рек.
В связи с этим задачами настоящей статьи и является сравнительный анализ: 1. Скорости и дальности
анемохорного и гидрохорного распространения семян сосны и ели. 2. Градиентов хорогенетической дифференциации популяций сосны обыкновенной на трансектах вдоль и поперек русел рек.
Объекты и методы
Изучение расселения самосева сосны от стены древостоя сосны обыкновенной высотой 28 м проведено на заброшенной пашне десятилетней давности в Припышминском лесном массиве подзоны предлесостепи Западной Сибири. Количество жизненного самосева 8-9-летнего возраста учтено на 37 учетных площадках размером 10 х 10 м, расположенных на трех трансектах на расстоянии от 50 до 1300 м от опушки леса.
Расселение самосева сосны обыкновенной и ели европейской выполнено в предгорьях Украинских Карпат на р. Ломнице, отличающейся бурным турбулентным течением. С этой целью на ее первой надпойменной террасе начиная с расстояния 150-200 м от локальных источников семян – культур сосны (в с. Сливки) и древостоя ели (в Ангеловском лесничестве) вплоть до лесного массива «Турова дача», расположенного на 17 км ниже по течению р. Ломницы, через каждые 1-4 км заложены пробные площади размером 20 х 100 м. На них проведен тщательный поиск и учет всех особей самосева сосны и ели. На протяжении изучавшегося участка реки другие источники семян, кроме исходных, не встречаются.
Плавательная способность семян сосны и ели изучена экспериментально. На поверхности воды в 10 стеклянных сосудах выкладывали по 100 семян с крылатками (имитация естественного налета семян) с определенной всхожестью (92%). Проводили ежедневный учет семян, оставшихся на плаву, и определение всхожести (по стандартной методике) утонувших семян на 3, 7, 11, 15, и 20-й дни после погружения на дно 90% семян.
Сравнение степени хорогенетической дифференциации и сходства смежных популяций сосны обыкновенной на трансектах в пределах речных бассейнов и на водоразделах между соседствующими бассейнами рек выполнено на основе результатов изозимного анализа (Корочкин и др., 1977) тканей почек 40-48 деревьев и определения генетических дистанций Неи (Nei, 1972, 1978).
Результаты и обсуждение
Анемохорное расселение самосева сосны. На рисунке 1 приведена кривая плотности жизненного самосева сосны обыкновенной 8-9-летнего возраста на равнинной поверхности заброшенной пашни после высокоурожайного семенного года (1997 г.). Численность самосева аппроксимируется уравнением экспоненты, уменьшаясь с 37,6 тыс. экз. на расстоянии 50 м до 0,25 тыс. экз. на расстоянии 1300 м от опушки соснового леса. Аналогичные данные о расселении самосева сосны обыкновенной на пашнях в южной тайге Русской равнины приводят А.П. Шиманюк (1955), а о распространении семян ели европейской метелью (по снежному насту) - Н.Е. Декатов 1961).
Можно предположить, что минимальная плотность древостоя, необходимая для перекрестного опыления и, следовательно, успешного продуцирования и дальнейшего "эстафетного" распространения семян сосны, составит 4-10 деревьев на 1 га, достигших репродуктивной стадии онтогенеза (как минимум 25-30-летнего возраста с хорошо развитой обильно семеносящей кроной). При среднем в данном регионе 20 - 30- процентном выживании самосева к этому возрасту его численность в 8 - 9- летнем возрасте должна быть не менее 15-20 экз./га. Экстраполируя данные по приведенному на рисунке 1 уравнению, находим, что такая плотность самосева может встречаться на расстоянии не далее 3000-3500 м от опушки (рисунок 1). 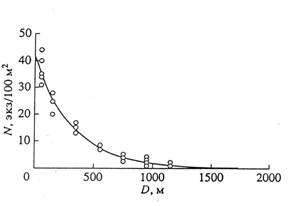
Рисунок 1 - Расселение самосева сосны обыкновенной на заброшенной пашне
D – расстояние от стены леса (м). N – плотность жизненного самосева 8-9-летнего возраста (экз./100 м2); N = 42,07exp (-0,0031D); R2 =0,97
Таким образом, в оптимальных ландшафтно-экотопических условиях возобновления популяций Pinussylvestris, в центре ее ареала, максимальная дальность достаточного расселения популяций сосны на идеально подготовленном, минерализованном субстрате пашни (аналог поверхности ледниковой морены) после исключительно урожайного семенного года может достигать не более 9-14 км за 100 лет, или 90-140 км в тысячелетие. В пессимальных климатических условиях перигляциальной (притундровой) зоны скорость продвижения популяций вслед за отступающим ледником должна быть намного меньше. Это обусловлено пониженной здесь в 3 – 5 раз семенной продуктивностью (Бойченко, 1970; Санников, 1992) и в несколько раз меньшей высотой и площадью форпостов древостоя.
Гидрохорное распространение семян и расселение самосева. Эксперименты, проведенные нами, показали, что семена сосны с крылатками на спокойной поверхности воды в сосудах могут находиться на плаву 5-8 дней (рисунок 2).
Позднее они постепенно тонут, но и через неделю после погружения на дно сохраняют довольно высокую всхожесть - в среднем 42%. При средней скорости течения равнинных рек около 2.5 км/час теоретически возможная дальность их распространения вдоль русла реки за 5 – 8 суток может достигать 300-480 км в год! Эта скорость на порядки величин выше темпа сухопутного анемохорного расселения популяций.
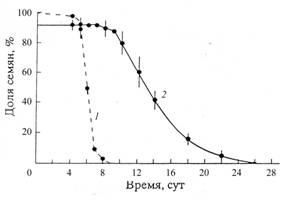
Рисунок 2 - Плавучесть и всхожесть семян сосны обыкновенной
1 – доля семян на плаву (%); 2 – всхожесть семян (%); Вертикальные линии – ошибки средних величин
Наши исследования на северо-восточном склоне Украинских Карпат в долине р. Ломницы, показали, что даже по течению бурных горных рек фактическая дальность гидрохорного распространения семян сосны обыкновенной и ели европейской составляет как минимум 17 – 20 км (рисунок 3). В годы наиболее высоких паводков (повторяющиеся через 10-12 лет) семена сосны и ели, разносимые речным течением, оседают на откосах первой надпойменной террасы, успешно прорастают и образуют жизненный самосев. За время до следующего паводка он обычно успевает достичь высоты 50-70 см, при которой его терминальные побеги уже надежно избегают затопления. Средняя плотность поселения самосева сосны на прирусловой террасе р. Ломницы на расстоянии 17 – 20 км от источников обсеменения составляет 40 – 45 экз./га, а ели – 20 – 23 экз./га. Таким образом происходит быстрое расселение популяций хвойных видов вдоль русла реки со скоростью не менее 60 – 100 км в столетие.
Можно предположить, что на равнинах Северной Евразии, где преобладает северное направление стока рек (за исключением южной половины Восточноевропейской равнины), преобладающим способом распространения семян главных лесообразующих хвойных из их южных рефугиумов (в Альпах, Карпатах, регионах Южного Урала, Казахского мелкосопочника и Южной Сибири) был и остается гидрохорный (Санников, Санникова, 2007). В неменьшей мере этот способ расселения присущ и лиственным видам древесных растений (особенно из родов Alnus, Corylus), а также семенам и раметам травянистых растений, вымываемым из обрывов береговых отложений (Удра, 1988).
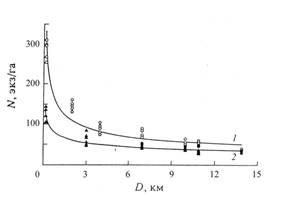
Рисунок 3 - Расселение самосева сосны (Pinussylvestris L.) и ели (Piceaabies Ledeb.) по течению реки Ломницы (Украинские Карпаты) от прибрежных источников семян:
1 – плотность самосева сосны N = 153,32D-0,43, R2 = 0,92; 2 – плотность самосева ели N = 78,61D -0,33, R2 = 0,76
Биогруппы и небольшие массивы взрослых деревьев, возникшие и выжившие в приречной зоне, становились форпостами дальнейшего быстрого гидрохорного распространения семян по течению рек, а также их сравнительно медленного анемо- и зоохорного расселения (вероятнее всего вихрями и птицами) вверх по прибрежным склонам и долинам притоков главной реки.
Генетическая интеграция популяций в бассейнах рек. Анализ генетических дистанций Неи (Nei, 1972, 1978) на протяжении 500 км вдоль русла р. Тобол и ее притоков показал (таблица), что между поселениями сосны на трансекте Кочердык–Курган– Заводоуспенское они в соответствии с нашей эмпирической шкалой (Санников, Петрова, 2003) не превышают ранг субпопуляции (0,005 - 0,006 по DN72 и 0,001-0,002 по DN78).
В бассейнах Днепра (Киев–Гомель) и Дона (Воронеж-Белая Калитва), Западной Двины (Архангельск–Сыктывкар) и Селенги (Кяхта–Улан-Удэ) генетические дистанции также обычно не превышают 0,008-0,010, не достигая уровня межпопуляционных различий.
По-видимому, группы популяций хвойных видов в рамках одного водосборного бассейна реки, как и микропопуляции животного населения, например мышевидных грызунов (Васильев, 1984), представляют некие линейные генетически слабо дифференцированные поселения, между которыми осуществляется постоянный гидрохорный нисходящий по долинам рек поток генов.
С другой стороны, для горных речных долин в часы дневной инсоляции характерны встречные восходящие потоки генов через пыльцу (Подгорный, 1981; Санников, Петрова, 2003). Это способствует формированию еще большей общности генофонда популяций древесных растений в пределах речного бассейна.
Генетическая дифференциация популяций между бассейнами рек. Анализ генетических дистанций Неи (Nei, 1972) между популяциями P. sylvestrisна трансектах, пересекающих водоразделы смежных речных бассейнов, например, Днепра и Дона, Тобола и Ишима, Селенги и Онона (таблица), а также Западной Двины и Волги, Волги и Камы, Оби и Енисея) показывает, что они в большинстве случаев находятся в пределах 0.013-0.026 (Санников, Петрова, 2003), т.е. в полтора-два больше, чем между популяциями, расположенными вдоль речного русла. При этом несмещенные дистанции Неи (Nei, 1978), которые между поселениями сосны, расположенными вдоль равнинных рек на расстоянии до 300-500 км, в большинстве случаев не превышают 0,002-0,004, на трансектах, пересекающих водоразделы, также достигают 0,009-0,011, т.е. популяционного уровня дифференциации. И, наконец, весьма значительные статистически достоверные генетические дистанции Неи (DN78 – 0,025-0,032) обнаружены (таблица) между горными группами популяций, расположенными в одном высотном поясе (650-850 м над уровнем моря) в смежных водосборных бассейнах рек Украинских Карпат Быстрица (Зеленская) и Прут (Яремчинская, Татаровская, Микуличин). В то же время между последними тремя выборками, находящимися в долине одной реки, DN78 примерно в два раза меньше (0,011-0,015). Возможно, это частью связано с высоким уровнем горно-механической изоляции анемохорных потоков семян и пыльцы между соседствующими долинами этих рек, разделенных отрогами хребтов с относительной высотой (над местообитаниями популяций) до 300-400 м. Однако, по-видимому не меньшее значение имеет постоянная противоположно направленная гидрохорная миграция семян сосны и, следовательно, гидрохорная изоляция популяций (Санников, 1993), расположенных в соседствующих водосборных бассейнах.Таким образом, имеются достаточные основания предполагать, что гидрохорная миграция семян и гидрохорная изоляция популяций - один из главнейших факторов, определящих степень хорогенетической интеграции и дифференциации популяций древесных растений.
Таблица. - Генетическая интеграция популяций сосны обыкновенной вдоль русел рек и дифференциация на трансектах поперек водоразделов
Наименование рек и водоразделов |
Популяции
(трансекты) |
Генетические дистанции (Nei,1972,1978) |
вдоль русла реки |
поперек водораздела |
DN72 |
DN78 |
DN72 |
DN78 |
Русская равнина |
Дон
Днепр
Дон-Днепр |
Воронеж- Б. Калитва
Гомель - Киев
Киев - Б. Калитва
Киев – Воронеж
Гомель – Б. Калитва |
0,005
0,008
-
-
- |
0,001
0,004
-
-
- |
-
-
0,014 |
-
-
0.011 |
Западная Сибирь |
Тобол
Тобол-Ишим |
Кочердык - Ик
Ик - Заводоуспенское
Кочердык-Орлиная гора
Заводоуспенское - Ишим
Ик – Орлиная гора |
0,005
0,006
-
-
- |
0,001
0,002
-
-
- |
-
-
0,013
0,013
0,013 |
-
-
0,010
0,009
0,010 |
Украинские Карпаты |
Прут
Прут-Быстрица |
Татаров – Микуличин
Яремча – Татаров
Яремча – Зеленская
Татаров- Зеленская
Микуличин – Зеленская |
0,015
0,019
-
-
- |
0.011
0,015
-
-
- |
-
-
0,035
0,029
0,029 |
-
-
0,032
0,026
0,025 |
Забайкалье |
Селенга
Онон
Селенга- Онон |
Улан-Удэ – Кяхта
Улан-Батор – Цасучей
Улан-Удэ – Цасучей
Кяхта – Цасучей |
0,007
0,016
-
- |
0,003
0,012
-
- |
-
-
0,026
0,015 |
-
-
0,023
0,012 |
Заключение
В итоге экспериментального и натурного изучения установлено, что максимальная скорость расселения самосева сосны обыкновенной в центре ее ареала (предлесостепь Западной Сибири) на оптимальном минерализованном субстрате заброшенных пашен не превышает 90-140 км /1000 лет, а скорость расселения самосева сосны и ели европейской по течению горной реки Ломницы (Украинские Карпаты) достигает 1700-2000 км/1000 лет, т.е. на порядок выше. Следовательно, гидрохория семян некоторых видов хвойных вдоль русел рек может быть одним из ведущих (возможно, приоритетных) факторов, определяющих пути и скорость миграции их популяций в плейстоцене (Санников, Санникова, 2007).
Сравнительный анализ межпопуляционных генетических дистанций Неи (Nei, 1978) показал, что на трансектах, ориентированных вдоль русел смежных рек Северной Евразии, они обычно не превышают 0,003-0,015 и в среднем в 1,5-2,5 раза меньше, чем на трансектах, расположенных поперек их водоразделов (0,009-0,032), особенно в горных регионах, где долины рек изолированы хребтами.
Таким образом, гидрохорная миграция семян – один из ведущих факторов, определяющих пути и степень геногеографической интеграции и дифференциации популяций древесных растений.
Библиографический список
- Бойченко, A.M.Произрастание сосны на северной границе ареала в Зауралье/ A.M.Бойченко // Экология. - 1970. - № 6. - С. 37 - 45.
- Васильев, А.Г.Изоляция расстоянием и дифференциация популяций/ А.Г.Васильев // Журн. общ. биол. 1984. - Т. 65.- №2. - С. 164 - 176.
- Гончаренко, Г.Г. Геосистематика и эволюционная филогения лесообразующих хвойных Палеарктики [текст]/ Г.Г. Гончаренко .- Минск: Технология, 1999.- 188 с.
- Декатов, Н.Е. Мероприятия по возобновлению леса при механизированных заготовках [текст]/ Н.Е. Декатов .- М. Л.: Гослесбумиздат, 1961. - 278 с.
- Корочкин, Л.И. Генетика изоферментов [текст]/ Л.И.Корочкин, О.Л.Серов, А.И.Пудовкин. - М.: Наука, 1977. - 275 с.
- Крутовский, К.В. Генетическая изменчивость сибирской кедровой сосны PinussibiricaDu Tour. Сообщ. 4. Генетическое разнообразие и степень генетической диффернциации между популяциями / К.В. Крутовский [и др.]// Генетика. - 1989. - Т. 25. - № 11. - С. 2009-2032.
- Ларионова, А.Я. Генетическое разнообразие и дифференциация популяций лиственницы Гмелина в Эвенкии (Средняя Сибирь) / А.Я. Ларионова // Генетика. -2004. - Т. 40. -№ 10. - С. 1370-1377.
- Петрова, И.В. Генетическая дифференциация болотных и суходольных популяций сосны обыкновенной в Западной Сибири / И.В. Петрова [и др.] // Экология. - 1989. - № 6. - С. 39-44.
- Потенко, В.В. Полиморфизм изоферментов и филогенетические взаимоотношения хвойных видов Дальнего Востока России: автореф. дис. … док. биол. наук./ В.В. Потенко.- Владивосток, 2004. - 38 с.
- Подгорный, Ю.К. О продолжительности периода рассеивания и фертильности пыльцы сосен / Ю.К. Подгорный // Науч. докл. высш. школы. Биол. науки, 1981.- № 1.- С. 74-78.
- Санников, С.Н. Возрастная биология сосны обыкновенной в Зауралье / С.Н. Санников // Восстановительная и возрастная динамика лесов на Урале и в Зауралье. Тр. Ин-та экологии растений и животных УНЦ АН СССР. - Свердловск, 1976. - Вып. 101. - С. 124-165.
- Санников, С.Н. Экология и география естественного возобновления сосны обыкновенной [текст]/ С.Н. Санников.- М.: Наука, - 1992. - 264 с.
- Санников,С.Н. Изоляция и типы границ популяций у сосны обыкновенной / С.Н. Санников // Экология. - 1993. - № 1. - С. 4 - 11.
- Санников, С.Н. Дифференциация популяций сосны обыкновенной [текст]/ С.Н. Санников, И.В. Петрова. - Екатеринбург: УрО РАН, 2003. – 248 с.
- Санников, С.Н. Гипотеза гидрохорного расселения популяций хвойных древесных растений / С.Н. Санников, Н.С. Санникова // Экология. - 2007. - № 2. - С. 83-87.
- Семериков, В.Л. Популяционная структура и молекулярная систематика видов LarixMill. автореф. дис. … док. биол. наук./ В.Л. Семериков.- Екатеринбург. - 2007. - 42 с.
- Удра, И.Ф. Расселение растений и вопросы палео- и биогеографии. [текст]/ И.Ф. Удра. - Киев: Наукова думка, 1988. - 196 с.
- Шиманюк, А.П. Естественное возобновление на концентрированных вырубках [текст]/ А.П. Шиманюк. - М: Изд-во АН СССР, 1955. - 355 с.
- Birks, H.J.В. Holocene isochrone maps and patterns of tree-spreading in the British Isles / H.J.В. Birks // J. Biogeogr. 1989. V. 16. P. 503 - 540.
- Cheddadi, R.. Imprints of glacial refugia in the modern genetic diversity of Pinus sylvestris / R.,Cheddadi // Global Ecol. Biogeogr., 2006. N. 15. P. 271-282.
- Firbas, F. Spat- und nacheiszeitliche Waldgeschichte Mitteleuropas nordlich der Alpen. Erster Band: Allgemeine Waldgeschichte / F. Firbas. Jena: Fischer, 1949. 480 S.
- Frencel, B. Grundzuge der pleistozanen Vegetationsgeschichte Nord-Eurasiens //Erdwissenschaft. Forschung. / B. Frencel. Wiesbaden: Steiner, 1968. Bd. 1. 326 S.
- Krutovskii, K. V., Bergmann F. Introgressive hybridization and phylogenetic relationships between Norway, Picea abies (L.) Karst., and Siberian, Picea obovata Ledeb., spruce species studied by isozyme loci / K. V. Krutovskii, , F. Bergmann // Heredity. - 1995. - V. 74. - P. 464-480.
- Lang, G.Quartare Vegetationsgeschichte Europas: Methoden und Ergebnisse / G.Lang,. Jena: Stuttgart; New York, 1994. 462 S.
- Lanner, R.M.Seed dispersal in Pinus II Ecology and biogeography of Pinus. Cambridge / R.M.Lanner, New York; Melbourne; Madrid: Cambridge University Press, 1998. P. 281 -295.
- Nei, M. Genetic distances between populations / M. Nei // Amer. Naturalist. 1972. V. 106. P. 283-292.
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals / M. Nei // Genetics, 1978. V. 89. P. 583 - 590.
- Prus-Glowacki. Allozyme differentiation in some European populations of Scots pine (Pinus sylvestris L.) / W.Prus-Glowacki, , L.Urbaniak, G. M. Zubrowska // Genetica Polonica. 1993. Vol. 34. № 2. P. 159-176.
- Semerikov, V.L. Intra- and interspecific allozyme variability in Eurasian Larix Mill. species / V.L.Semerikov, , L.F.Semerikov, M. Lascoux // Heredity. 1999. Vol. 83. P. 193-204.
- Yazdani, R., Muona O., Rudin D., Szmidt A.E. Genetic structure of a Pinus sylvestris L. seed-tree stand and naturally regenerated understory / R. Yazdani, , O. Muona, D. Rudin, A.E. Szmidt // Forest Sci.. Vol. 31. № 2. P. 430-436.
|


