|
"Хвойные бореальной зоны" 2007г.,№2-3, с.
СОХРАНЕНИЕ И УЛУЧШЕНИЕ ГЕНЕТИКО-СЕЛЕКЦИОННОГО ПОТЕНЦИАЛА ОСНОВНЫХ ЛЕСООБРАЗУЮЩИХ ВИДОВ В УСЛОВИЯХ СРЕДНЕГО УРАЛА
Махнев А.К., Лебедев Ю.В.*, Уманова Н.Е., Лебедев А.Е.*
Ботанический сад Уральского отделения РАН, Екатеринбург, Россия
620134, г. Екатеринбург, ул. Балимбаевская, 32а, т. (343) 3225629, afrmah@rambler.ru
*Сысертская лесная производственная семеноводческая станция, Сысерть, Россия
Исследованы испытательные культуры (ИК) первого порядка Pinussylvestris L. и Piceaobovata Ledeb., заложенные в 1981-1997 гг. на юге Свердловской области в географическом градиенте, включающем пункты, расположенные в Предуралье, Горном Урале и Зауралье. Целью работы являлась сравнительная генетико-селекционная оценка потомств плюсовых деревьев в ИК для идентификации элитных деревьев, перспективных для закладки ИК второго порядка. В качестве материала для соответствующей оценки использованы результаты периодической регистрации роста и состояния потомств 374 плюсовых деревьев. Оценка структуры ИК по энергии роста и состоянию с помощью трехфакторного иерархического и других моделей дисперсионного (однофакторного, двухфакторного) анализов позволила выявить долю влияния отдельных факторов или вклад их в общую дисперсию. При этом установлено, что доля влияния фактора географического происхождения плюсовых деревьев, составляющая в среднем лишь несколько процентов, минимальна, но в отдельных опытах статистически значима. Далее по вкладу, обычно не превышающему 14 %, следует фактор семейной принадлежности. Затем следует фактор повторности, включающий взаимодействие генотип-среда, вклад которого составляет около 20 %. Завершает данный ряд факторов остаточная дисперсия, приходящаяся на индивидуальную изменчивость, доля которой доходит до 90 %. Таким образом, установлено, что даже на довольно ограниченной в географическом плане территории сформировались популяции лесообразующих видов, в т.ч. сосны обыкновенной, скоррелированные по своим эколого-биологическим свойствам со своеобразным природно-климатическим комплексом, что отражается на энергии роста потомств плюсовых деревьев из этих популяций. Данный феномен свидетельствует о перспективности популяционной селекции, наравне с индивидуальной.
The first order (generation) test cultures of a Pinus sylvestris and Picea obovata, layed up in 1981-1997 yy. in the south of Sverdlovsk region in a geographical gradient including points, arranged in the Western Ural, in the Mountain Ural and in the Eastern Ural are investigated. The purpose of work was the comparative genetic-selection estimation of descendants of plus trees in test cultures for identification of elite trees, perspective for laying of the test cultures second order. As a material for an appropriate estimation the outcomes of periodic registration of growth and state of descendants of 374 plus trees are used. The estimation of frame test cultures on energy of growth and state with the help three-factor hierarchic and other models dispersion (one-factor, two-factor) analyses has allowed to reveal a share of effect of the separate factors or contribution them in a general dispersion. At that established, that the share of effect of the factor of a geographical parentage of plus trees make up on the average some of percents, is minimum, but in separate experience is statistically significant. Further under the contribution, 14 % is customary not superior the factor of a family fitting follows. Then the factor repetitive experiment, including interaction genotype - environment, follows, the contribution makes which one about 20 %. Completes the given number of the factors a residual dispersion, coming on individual variability, the share by which one reaches 90 %. Thus, established, what even on territory, rather limited in the geographical schedule, the populations forestformes of species were formed, including Pinus sylvestris, correlated on the ecologycal-biological properties with a peculiar nature-climatic complex, that is mirrored in energy of growth of descendants of plus trees from these populations. The given phenomenon testifies to prospects of population selection, level with individual.
Как известно, генетико-селекционный потенциал видов представлен разными уровнями, что следует из политипической концепции, согласно которой на верхних ступенях внутривидовой иерархии существуют ареальные единицы, имеющие таксономический ранг, например, подвид (разновидность). В то же время внутривидовая структура включает ряд ареальных единиц, в том числе достаточно обширных, не имеющих таксономического ранга – экотип, климатип, группа популяций, популяция, микропопуляция и их различные варианты (Правдин, 1964, 1965; Мамаев, 1970; Скворцов, 1974 и др.). Примечательно, что в рамках фундаментальных исследований второй половины прошлого века, посвященных изучению закономерностей внутривидовой изменчивости и формирования популяционной структуры, в т.ч. основных лесообразующих видов, были экспериментально установлены два основополагающих момента: 1) наличие строгой структурированности в виде иерархических рядов популяций разного ранга; 2) тесная взаимосвязь и взаимодействие данной структуры с природно-климатическим комплексом (Мамаев, 1972; Семериков, 1986; Махнев, 1987), что в сущности означает взаимодействие двух систем интеграции – популяционно-видовой и биогеоценотической (Шварц, 1980). Немаловажный интерес в генетико-селекционном отношении также представляет оценка структуры внутривидовой изменчивости с точки зрения вклада ее отдельных форм (уровней) в общую дисперсию, показавшая, что наибольший вклад в данном случае приходится на долю индивидуальной изменчивости, а далее межпопуляционной (Глотов и др., 1975; Махнев, 1987). Более поздние генетические исследования, выполненные с помощью изоферментного анализа и изучения ДНК (Духарев, Правдин, 1983; Янбаев, 2002; Санников, Петрова, 2003), также убедительно показали, что наряду с индивидуальным отбором может быть достаточно эффективной и популяционная селекция. Последнее уже неоднократно показано благодаря наличию географических и популяционно-экологических культур, созданных во многих странах мира еще в конце XIX века (Ромедер Э., Шёнбах Г., 1962; Райт Дж. В., 1978 и др.), в т.ч. в России, хотя наиболее интенсивно в этом направлении исследования проводились уже во второй половине XX века.
при создании единого селекционно-генетического комплекса исключительно важное значение имеет корректный выбор соответствующей стратегии. Практически априори в основу была положена так называемая плюсовая селекция, базирующаяся на индивидуальном отборе. Однако уже достаточно продолжительный опыт показал, что результаты ее во многом противоречивы, что связано с наличием явных плюсов и минусов, что также отмечено в работе А.С. Бондаренко (2002).
к первому относятся такие моменты как возможность получения качественного материала (семян и черенков) с лесосеменных плантаций (ЛСП) и достаточно высокое генетическое разнообразие этих искусственных популяций, позволяющие применять интенсивные методы селекции, например, селекцию на гетерозисный эффект. Кроме того, индивидуальный отбор – это четкая идентификация генотипов и поэтому однозначная воспроизводимость материала на основе вегетативного размножения. Высокая интенсивность индивидуального отбора также позволяет достигнуть необходимого селекционного эффекта. Так, обобщение опыта создания ЛСП показало, что генетический эффект ЛСП первого порядка, определяемый по быстроте роста испытательных культур (ИК), оценивается в среднем на уровне 8-9% с амплитудой 5-25% (Бондаренко, 2002).
К числу основных недостатков принятой в России схемы плюсовой селекции следует, очевидно, отнести трудность идентификации генотипа по фенотипу, что влечет за собой сложность и длительность оценки потомств в ИК. Кроме того, ограниченное число генотипов, используемое при данной схеме, и направленный искусственный отбор ведет к снижению генетического разнообразия. Тем не менее, пока однозначная оценка упомянутой схемы селекции отсутствует, работы по ее реализации с критической оценкой активно продолжаются.
Одним из крупных объектов в этом плане являются ИК плюсовых деревьев, созданные Сысертской лесной производственной семеноводческой станцией (ЛПСС) в рамках обширной программы, посвященной созданию в лесхозах Свердловской области единого генетико-селекционного комплекса (ЕГСК).
Целью нашего исследования была сравнительная генетико-селекционная оценка потомств плюсовых деревьев в ИК первого порядка сосны обыкновенной (Pinussylvestris L.) и ели сибирской (Piceaobovata Ledeb.), заложенных на юге Свердловской области в 1981-1997 гг. в географическом градиенте, включающем пункты, расположенные в Предуралье, Горном Урале и Зауралье, для идентификации элитных деревьев. В задачи исследований входило: произвести сравнительный анализ полевых материалов и дать корректную оценку перспективности использования отдельных семей в качестве исходных для закладки лесосеменных плантаций второго порядка, а также разработать рекомендации по их закладке на генетико-селекционной основе.
Объекты и методы исследований
Объектом исследований являются ИК первого порядка сосны обыкновенной и ели сибирской, заложенные в Свердловской области Сысертской ЛПСС в различных лесорастительных условиях (Колесников, 1969): на территории Сысертского, Тугулымского, Ачитского лесхозов и национального природного парка (НПП) «Припышминские боры» (Талицкий и Тугулымский лесхозы), которые занимают 20,75 га (20,15 га сосны и 0,6 га ели) (Лебедев, Заровнятных, 2000). Испытывались потомства плюсовых деревьев шести происхождений сосны обыкновенной: Сысертского, В-Исетского и Асбестовского (Средний Урал), Камышловского (Средний Урал – Зауралье), Талицкого и Тугулымского (Зауралье); одного ели сибирской: Красноуфимского (Предуралье). В ИК, созданных в период с 1981 по 1997 год, прошедших соответствующую оценку в порядке мониторинга, испытывается потомство 374 плюсовых деревьев. Из них в ИК разных лет представлены: один раз – 242 плюсовых дерева, два раза – 124 плюсовых дерева, три раза – 8 плюсовых деревьев. Основная часть ИК в Свердловской области, составляющая 79,3 % от общей площади, заложена на территории Сысертского лесхоза. Вторым по занимаемой площади ИК сосны обыкновенной является Тугулымский лесхоз, где в течение 3-х лет (1987, 1988 и 1996 гг.) они были созданы на площади 2,4 га.
Согласно лесорастительному районированию Свердловской области (Колесников, 1969) территория, на которой расположен основной массив ИК сосны обыкновенной, относится к Западно-Сибирской равнинной лесной области, Зауральской холмисто-предгорной провинции сосново-березовых предлесостепных лесов. По двум из основных экологических факторов - теплу и осадкам - две сравниваемые группы районов – Ачитский и Сысертский, с одной стороны, и Талицкий и Тугулымский, с другой стороны, существенно отличаются, что не сможет не сказаться на других показателях, например, эдафических, характеризующих плодородие почв (таблица 1). Для обеспечения получения достоверных данных на всех этапах оценки (краткосрочном, среднесрочном и долгосрочном) все работы выполнялись в соответствии с Основными положениями методики закладки ИК плюсовых деревьев главных лесообразующих пород (1979). В соответствии с этим схема опытов включает: определенное число вариантов (семей), число растений на одной делянке, составляющее 10 деревьев и повторность, т.е. число блоков (не менее четырех). В числе вариантов также присутствует контроль, представляющий средний образец из семян производственного сбора.
По всем годам закладки ИК выдержан основной принцип размещения делянок на экспериментальной площади, так называемая рандомизация. При посадке опытные растения высаживали рядами с размещением 3x1 м.
В связи с одной из основных целей селекции – повышением продуктивности и качества лесных насаждений (древостоя) главных лесообразующих видов - в процессе мониторинга ИК изучались и оценивались на первом этапе прежде всего быстрота роста (по высоте и диаметру), качество ствола и устойчивость к экстремальным факторам среды. Обработка всех материалов, полученных в результате мониторинга ИК, производилась на персональном компьютере в ПСП STATISTICA 5.5 (Statsoft, Inc.2001). В качестве основного метода обработки для последовательного решения поставленных задач использовался дисперсионный анализ: однофакторный, иерархический многофакторный и двухфакторный (Рокицкий, 1973).
На первом этапе проверялась степень однородности ряда, представленного разными происхождениями семей в посадках одного года с помощью как однофакторного, так и иерархического трехфакторного анализов. В последнем случае также оценивалась доля вклада отдельных компонентов внутривидовой структуры в общую дисперсию: разных географических происхождений (природных популяций), семей (потомств плюсовых деревьев в культуре), повторностей, а также отдельных деревьев (индивидов), характеризующих долю индивидуальной изменчивости.
На втором этапе проводился двухфакторный дисперсионный анализ (факторы семья и повторность) с учетом контроля (условно принимался как семья), в результате которого, главным образом, выявлялось наличие или отсутствие достоверных различий между семьями, включая контроль. В случае установленного на первом этапе статистически значимого различия географических происхождений анализ проводился отдельно для каждого из них.
На третьем этапе проводилось ранжирование семей каждого года испытаний по каждому году измерений, выявлялись семьи, превышающие контроль на 15-20% и, в случае ранее выявленной значимости влияния фактора семьи, проверялась достоверность различий между потомствами лучших семей и контролем. Различия считались значимыми на 95-99% и выше (p<0,05; p<0,01) при попарных сравнениях по Т-критерию Стьюдента и по критерию множественных сравнений при однофакторном дисперсионном анализе (HSD Tukey test). Тест множественных сравнений Тьюки для неравных размеров выборок из предлагаемого программой STATISTICA проверочных критериев был выбран как наиболее корректный применительно к анализируемому материалу.
Четвертым этапом была идентификация плюсовых деревьев, семьи (потомство) которых на 15-20% стабильно как в хронологическом ряду, так и при множественных испытаниях значимо превосходило контроль. По результатам комплексной оценки лучшие плюсовые деревья при краткосрочной оценке рекомендовались в условные кандидаты в плюсовые элитные деревья и при среднесрочной оценке в кандидаты в плюсовые элитные деревья.
Данный последовательный анализ позволяет всесторонне и корректно определить выдающиеся плюсовые деревья испытываемых в культурах лесообразующих видов, частично ответить на вопрос о целесообразности и пределах переброски улучшенных (элитных) семян сосны Свердловской области из одних природно-климатических районов в другие, поскольку сосна на данных территориях (районах) отличается на уровне групп популяций (Мамаев, 1970); расширить представления о целесообразности повторных закладок ИК, используя семена одних и тех же плюсовых деревьев, но урожаев разных лет, а также по вопросу, касающемуся возраста ИК, и получить удовлетворительный ответ относительно селекционно-генетической основы их наследственных особенностей.
Экспериментальная часть
Анализ структуры изменчивости, выполненный с помощью трехфакторного дисперсионного анализа (неортогональный комплекс), позволил оценить соотношение паратипических (повторность) и генетических (происхождение, семья и индивидуальная) компонент изменчивости.
Краткосрочная оценка на этапах 5 и 10 лет (таблица 2) показывает, что основная доля дисперсии (от 66,4 до 88,4 %) приходится на индивидуальную изменчивость. Далее, как правило, на статистически значимом уровне представлен в рассматриваемой структуре изменчивости уже паратипический компонент – повторность. Затем, более чем в 50% случаев статистически значим вклад в общую дисперсию семей отдельных плюсовых деревьев, хотя в ИК 1981 и, частично, 1994 года он весьма незначителен (от 0,2 до 2,4 %). Вклад происхождения (географического) в общую дисперсию в большинстве случаев незначителен или вообще отсутствует, но в посадках 1981 года по оценке в 5-летнем возрасте оказался значим. Однако обращает на себя внимание то, что в ИК сосны, заложенных в 1997 году, соответствующий вклад оказался весьма существенным и статистически значимым – 14,5 %.
Таким образом, в краткосрочном аспекте фиксируются три основных момента: существенное повышение вклада в общую дисперсию в ряду географическое происхождение-семья-особь (1); увеличение вклада фактора семейной принадлежности в общую дисперсию с возрастом (2) и, напротив, снижение на уровне происхождения (3) (таблица 2).
В целом, несмотря на довольно небольшой вклад географического происхождения в рамках рассматриваемых районов в общую дисперсию, свойственную (присущую) ИК сосны, определенную в системе трехфакторного иерархического комплекса, по данным однофакторного дисперсионного анализа географические происхождения, особенно с крайних пределов рассматриваемых рядов популяций, как правило, существенно различаются, в том числе по энергии роста.
Таблица 1 – Основные сведения о климате (по данным Агроклиматического справочника по Свердловской области, 1962) в районе участков ИК
Административный
район |
Продолжительность периода, дни |
Сумма положительных температур выше 10 ?С |
Сумма осадков, мм (за год / за период с температурой воздуха выше 10 ?С) |
Гидротермический показатель |
Продолжительность периода с устойчивым снежным покровом, дни |
Средняя высота снежного покрова, см |
безморозного |
с температурой воздуха выше 10 ?С |
Ачитский |
90-95 |
115 |
1600-1750 |
550 / 250 |
1,4-1,6 |
165-170 |
60 |
Сысертский |
100-105 |
115 |
1600-1800 |
450 / 250 |
1,4-1,6 |
150-155 |
45 |
Талицкий |
105-110 |
125 |
1850-1900 |
375 / 225 |
1,2 |
145-150 |
45 |
Тугулымский |
95-105 |
125 |
1825-1875 |
425 / 225 |
1,2 |
150-155 |
40 |
Необходимо отметить, что по данным однофакторного дисперсионного анализа ИК 1995 г., заложенных в Сысертском лесхозе, не наблюдается достоверных различий между среднеуральскими (Сысертский, Асбестовский) и зауральским климатипами (Талицкий) при тенденции снижения показателя роста, но более высокого уровня внутрипопуляционной изменчивости последнего (рисунок 1). В ИК сосны 1996 года, представленных максимальным числом географических происхождений: Сысертским, Свердловским, Верх-Исетским, Асбестовским, Талицким и Тугулымским, последний статистически значимо отличается от всех остальных сравнительно низким показателем роста по высоте. Кроме него, от остальных, за исключением Сысертского климатипа отличается Асбестовский. В ИК сосны 1997 года, заложенных в Сысертском лесхозе, Тугулымский климатип также в сравнении с Сысертским, Асбестовским и Талицким показал достоверно самый низкий результат. В связи с этим примечателен такой факт, что в ИК сосны данного года, но заложенных в Тугулымском лесхозе, достоверные различия между Сысертским, Тугулымским и Асбестовским географическими происхождениями по энергии роста отсутствуют (рисунок 1).
По данным двухфакторного дисперсионного анализа с учетом контроля при краткосрочной оценке дифференциация на межсемейном уровне обнаружена практически у всех ИК, за исключением ИК 1981 г. Сысертского (11 семей) и Камышловского (4 семьи) происхождений в Сысертском лесхозе, в то время как в наиболее репрезентативном Талицком происхождении (14 семей) все три компонента дисперсии (семья, повторность, взаимодействие генотип-среда) оказались значимыми. При этом взаимодействие факторов семьи и повторности оказалось статистически значимым практически во всех ИК, а в ИК 1997 г. в Сысертском лесхозе Сысертского (33 семьи) и Тугулымского (26 семей) происхождений оказался значим фактор повторности, хотя в Тугулымском лесхозе в ИК 1997 г. при учете всех происхождений (51 семья) он не оказался существенным.
В анализируемых ИК среднесрочная оценка их энергии роста, развития и состояния охватывает период 5-20 лет. В данные временные рамки включаются ИК, заложенные в 1982, 1983, 1984, 1987, 1988 и 1990 годах в Сысертском лесхозе и в 1988 году в НПП «Припышминские боры».
Данные таблицы 2, в которой по результатам трехфакторного иерархического анализа оценивается доля вклада различных факторов (географического происхождения, принадлежности к семье, повторности, индивидуальной принадлежности) в общую дисперсию, убедительно свидетельствуют о том, что в ИК 1982 года на фоне очень высокой доли индивидуальной изменчивости (76, 7-91,03 %) вклад географического происхождения практически равен нулю или очень незначителен. Вклад фактора семьи в общую дисперсию по высоте становится значимым к 15-летнему возрасту. Однако к 20-летнему, в связи с очень высокой изменчивостью по диаметру, нивелируется. Вклад повторности оказался сравнительно высоким на всех этапах оценки (за исключением диаметра по указанной причине). В ИК 1983 и 1984 годов закладки только материалом Сысертского происхождения существенным оказался вклад в общую дисперсию фактора принадлежности к семье по высоте, но по диаметру по указанной выше причине этого не наблюдалось. Вклад фактора повторности оказался существенным по среднесрочной оценке из рассматриваемых только в ИК 1984 года, хотя в возрастном ряду на начальном этапе роста он был значим и в культурах 1983 года. В ИК 1987 года, как и 1988 года, заложенных в Сысертском лесхозе, на фоне очень высокой индивидуальной изменчивости диаметра в 15-летнем возрасте вклад всех рассматриваемых факторов, за исключением индивидуальной изменчивости, оказался несущественным. Однако в ИК 1988 года, заложенных в НПП «Припышминские боры», и 1990 года, заложенных в Сысертском лесхозе, где вклад фактора остаточной (индивидуальной) изменчивости оказался менее существенным, статистически значимым является вклад в общую дисперсию фактора повторности на уровне 6,9-12,3%. Однофакторный дисперсионный анализ (рисунок 2) в значительной степени позволил подтвердить данные иерархического дисперсионного анализа, поскольку фактор географического происхождения оказался значим только в культурах 1990 года, в том числе по высоте. В данном случае Сысертское происхождение отличается от Верх-Исетского и Талицкого, а Асбестовское от Талицкого (по Т-критерию). По диаметру статистически значимо отличается только Талицкое происхождение, представленное одной семьей. Необходимо также отметить тенденцию более высоких показателей по диаметру Сысертского происхождения в сравнительном плане между происхождениями ИК 1988 г. в Сысертском лесхозе, по сравнению с ИК этого же года в НПП “Припышминские боры” (рисунок 2). Результаты двухфакторного дисперсионного анализа показателей роста ИК сосны с учетом контроля при среднесрочной оценке во временной динамике свидетельствуют о том, что, по крайней мере, в 50% рассматриваемых случаев дифференциация семей, в отличие от таковой по географическим происхождениям, оказалась значимой. В частности, в ИК, заложенных в 1982 году, лишь в 5-летнем возрасте дифференциация семей по высоте не оказалась существенной. Далее в 10-ти и 15-ти летнем возрасте по высоте и в 20-летнем по диаметру она была достоверной, хотя влияние среды (повторность) за данный период оказалось недостоверным дважды (в 10-ти и 20-летнем возрасте), а взаимодействие факторов «генотип-среда» только при мониторинге в 20-летнем возрасте. Своеобразная динамика дифференциации по энергии роста наблюдается в ИК, заложенных в 1983 году в Сысертском лесхозе. В данном случае в 4-летнем возрасте их дифференциация культур на уровне всех анализируемых факторов оказалась существенной, но при мониторинге в 10-летнем возрасте она оказалась достоверной только на межсемейном уровне. При мониторинге в 20-летнем возрасте она не была зафиксирована ни по одному из анализируемых факторов.
По результатам мониторинга посадок сосны 1984 г. в 5-летнем возрасте дифференциация оказалась существенной на семейном уровне и на уровне взаимодействия «генотип-среда». Однако по результатам мониторинга данных культур в 20-летнем возрасте по диаметру наличие дифференциации на межсемейном уровне не подтвердилось.
Своеобразным исключением в отношении степени дифференциации практически одновременно по всем анализируемым показателям (факторам) являются ИК, заложенные в 1987 году в Сысертском лесхозе. У данных культур, представленных Сысертским, Камышловским и Талицким происхождениями, по результатам мониторинга в 5-ти и 15-ти летнем возрасте достоверная дифференциация не выражена ни по одному фактору, а в 10-ти летнем возрасте выражена только по взаимодействию факторов «генотип-среда». Аналогичная с посадками 1987 года картина наблюдается также в посадках 1988 года, произведенных в Сысертском лесхозе. Однако в ИК, заложенных в данном году в НПП «Припышминские боры», достоверная дифференциация выражена по фактору среды и взаимодействию «генотип-среда».
Напротив, в ИК, заложенных в 1990 году, дифференциация, судя по результатам мониторинга в 10-ти и 15-ти летнем возрасте, стабильно выражена практически по всем анализируемым факторам (за исключением взаимодействия факторов «генотип-среда» в 15-летнем возрасте).
Таким образом, в среднесрочном аспекте, как и в краткосрочном, также фиксируется повышение вклада в общую дисперсию в ряду географическое происхождение – семья - особь, однако тенденция изменения с возрастом вклада семейной принадлежности в определенной степени завуалирована, так как с возрастом относительно стабильный показатель – высота заменен на более лабильный – диаметр.
При решении основной задачи работы – идентификации кандидатов в элитные плюсовые деревья сосны обыкновенной целесообразно изложить основные моменты, показывающие, что формальный подход, позволяющий относить к таковым те, которые только превышают контроль по энергии роста на 15-20%, не вполне корректен по ряду очевидных причин, основные из которых следующие:
1. Подавляющее большинство плюсовых деревьев не прошло повторную (двукратную) проверку в ИК с использованием для этого семян урожая разных лет.
2. Представительность контроля в ИК, находящаяся в пределах 1,5-6,7% (в среднем 2,5%), достаточно низкая.
3. Изменение оценочных параметров в возрастном ряду, например, высоты на диаметр.
4. Наличие существенных отклонений по представительности деревьев в пределах отдельных повторностей в ИК.
Учитывая наличие этих моментов, негативно влияющих на корректность определения (отбора) кандидатов в элитные плюсовые деревья в число оценочных тестов, как и при соответствующей идентификации ели сибирской, при комплексной оценке использованы критерии: множественных сравнений, рассчитываемый при однофакторном дисперсионном анализе между семьями с учетом контроля (Tukey HSD тест), T-критерий, наличие превышения контроля по оцениваемым параметрам на 15 %, результаты повторных испытаний на отдельных этапах и испытаний в разных лесорастительных условиях.
Таким образом, в результате комплексного анализа материалов мониторинга ИК сосны обыкновенной, заложенных в период 1981-1997 гг. в нескольких лесхозах Свердловской области, идентифицировано 50 плюсовых деревьев, которые согласно существующим нормативным документам и имеющихся рабочих проектов могут быть использованы как база для создания ИК второго порядка и лесосеменных плантаций.
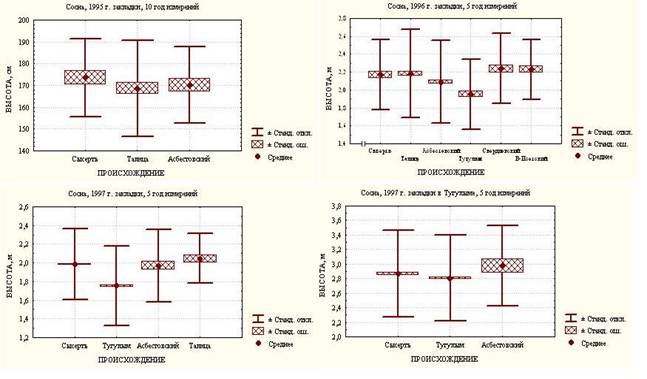
Рисунок 1 – Показатели роста сосны в ИК Сысертского и Тугулымского лесхозов при краткосрочной оценке с учетом происхождения
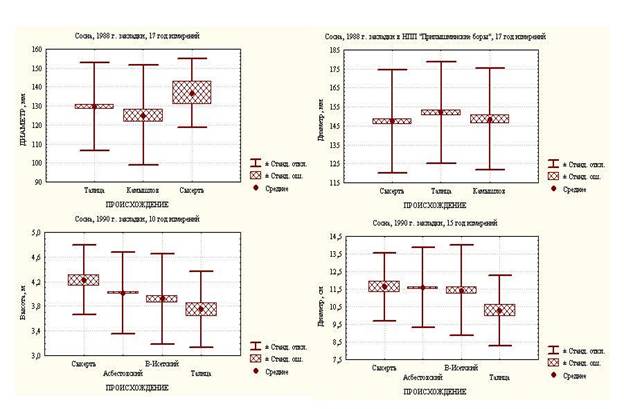
Рисунок 2 – Показатели роста сосны в ИК Сысертского лесхоза и НПП “Припышминские боры” при среднесрочной оценке (1988, 1990 гг.) с учетом происхождения
Учитывая продолжительность периода испытаний и стабильность показанных результатов, а, следовательно, и корректность оценки наследственных свойств отобранных деревьев, они подразделяются на две категории:
- условные кандидаты в элитные плюсовые деревья, т.е. в основном прошедшие краткосрочную оценку;
- кандидаты в элитные плюсовые деревья, т.е. прошедшие среднесрочную оценку.
В число первых включено 24, а вторых – 26 плюсовых деревьев сосны обыкновенной.
ИК ели сибирской заложены в 1990 году в Ачитском лесхозе на площади 0,6 га и представлены семьями 30 плюсовых деревьев, отобранных в Нижне-Сергинском и Ачитском лесничествах Ачитского лесхоза. Двухфакторный дисперсионный анализ, включающий контроль, при среднесрочной оценке ИК ели показал статистически значимое влияние на энергию роста всех рассматриваемых факторов. Основная доля дисперсии 79,3% приходится на остаточную (индивидуальную) изменчивость. Далее в порядке снижения следует фактор взаимодействия «генотип-среда» с вкладом в общую дисперсию доли, составляющей 10,7%, несколько меньше доля, характеризующая межсемейную изменчивость, – 7,7% и самая минимальная доля приходится на фактор, отражающий влияние повторностей. Указанное соотношение вклада перечисленных факторов в общую дисперсию, характерных для ИК ели сибирской, в общих чертах соответствует таковому, установленному в ИК сосны обыкновенной.
В связи с отсутствием промежуточных этапов мониторинга ели, в возрасте 5 и 10 лет, больших (83,3 %) семей, превышающих контроль на 15 % по высоте, и рядом причин, ранее указанных для сосны, отбор кандидатов в элитные плюсовые деревья ели произведен более жестко, в т.ч. с использованием Tukey HSD теста. В результате в число кандидатов в элитные деревья попали 11 плюсовых деревьев, которые в соответствующем ранжированном ряду заняли первые места, т.е. фактически превышающие контроль по высоте более, чем на 28,2% и статистически значимо отличающиеся от него по энергии роста по указанному критерию.
В результате проведенных исследований сделаны рекомендации по закладке лесосеменных плантаций второго порядка на генетико-селекционной основе, которые, прежде всего, касаются перечня идентифицированных в результате использования комплекса тестов кандидатов и условных кандидатов в элитные плюсовые деревья сосны обыкновенной и ели сибирской. Важным аспектом является необходимость увеличения представительности и сбалансированности контроля с целью организации ортогональных комплексов, более корректных при математико-статистической оценке энергии роста и других показателей ИК. Далее, учитывая наличие значительного влияния на корректность сравнительной оценки показателей роста на межсемейном уровне факторов, связанных со степенью выравненности условий среды и, соответственно, взаимодействия генотип-среда следует обеспечить относительную однородность фона в первую очередь по эдафическим условиям.
Кроме того, учитывая положительные и отрицательные моменты популяционной и плюсовой селекции, базирующихся соответственно на массовом и индивидуальном отборе, целесообразно обеспечить сбалансированное сочетание этих двух наиболее апробированных и эффективных в лесохозяйственном производстве направлений селекции, что обеспечит сохранность биоразнообразия основных лесообразующих видов на популяционном уровне и повышение продуктивности и устойчивости их древостоев.
Заключение
Анализ ИК, представленных потомствами плюсовых деревьев разного географического происхождения, позволил оценить долю вклада отдельных факторов в общую дисперсию показателей энергии роста. В целом установлено, что соответствующая доля фактора географического происхождения плюсовых деревьев, составляющая в среднем лишь несколько процентов, минимальная, далее по вкладу обычно не превышающему, или незначительно превышающему 10%, следует фактор семейной принадлежности. Затем следует фактор повторности, включающий также взаимодействие генотип-среда, вклад которого находится в пределах или несколько больше 20%. Завершает данный ряд факторов остаточная дисперсия, приходящаяся на индивидуальную изменчивость, доля которой может составлять более 90%.
Хотя вклад географического происхождения в общую дисперсию сравнительно мал, и в отдельных случаях статистически не значим, там не менее даже на довольно ограниченной в географическом плане территории, как, например, южные пределы Свердловской области в широтном направлении, сформировались популяции лесообразующих видов, в том числе сосны обыкновенной, скоррелированные по своим эколого-биологическим свойствам со своеобразным природно-климатическим комплексом, что находит отражение на энергии роста потомств плюсовых деревьев из этих популяций, особенно наиболее удаленных от района закладки ИК. Данный феномен убедительно свидетельствует о перспективности и приоритете популяционной селекции, в том числе сосны обыкновенной, произрастающей в условиях достаточно контрастных по природно-климатическим особенностям фона.
Благодарности
Авторы статьи выражают благодарность сотрудникам Сысертской лесной производственной станции Т.И. Заровнятных и Л.П. Светлаковой, выполнявших систематический мониторинг по оценке энергии роста и состояния ИК.
Таблица 2 – Структура изменчивости высоты испытательных культур сосны при краткосрочной оценке по результатам иерархического трехфакторного дисперсионного анализа
Год закладки, лесхоз |
Доля вклада фактора в общую дисперсию, % |
1981, Сысертский |
1994, Сысертский |
1995, Сысертский |
1996, Сысертский |
1997, Сысертский |
1997, Тугулымский |
Год измерений |
5 |
10 |
5 |
10 |
5 |
10 |
5 |
5 |
5 |
Происхождение |
2,8* |
1,5 |
1,3 |
0 |
0 |
0 |
0 |
14,5*** |
0 |
Семья |
1,2 |
0,2 |
2,4 |
4,0* |
14 |
9,8*** |
13,6*** |
3,8** |
10,9*** |
Повторность |
21*** |
23*** |
7,8*** |
8,5*** |
- |
12,2 |
21,5*** |
15,3*** |
13,1*** |
Остаточная |
75,0 |
75,3 |
88,5 |
87,5 |
86,0 |
77,9 |
64,9 |
66,4 |
76,0 |
Примечание - * p<0,05, ** p<0,01, *** p<0,001. |
Таблица 3 – Структура изменчивости ростовых параметров испытательных культур сосны при среднесрочной оценке по результатам
иерархического трехфакторного дисперсионного анализа при закладке в Сысертском лесхозе и в 1988 г. в НПП «Припышминские боры»
Год закладки |
Доля вклада фактора в общую дисперсию, % |
1982 |
1983 |
1984 |
1987 |
1988 |
1988НПП |
1990 |
Год измерений |
5 |
10 |
15 |
20 |
4 |
10 |
20 |
5 |
20 |
5 |
10 |
15 |
15 |
17 |
10 |
17 |
Параметр |
высота |
высота |
высота |
диаметр |
высота |
высота |
диаметр |
высота |
диаметр |
высота |
высота |
диаметр |
диаметр |
диаметр |
высота |
диаметр |
Происхождение |
0 |
0 |
0 |
0,09 |
|
|
|
|
|
0 |
0,40 |
0 |
0,43 |
0 |
0 |
0,71 |
Семья |
2,9 |
5,3 |
10,4** |
1,68 |
5,7** |
23,4** |
0 |
6,8** |
0 |
0 |
0 |
0 |
1,0 |
0 |
2,74 |
0,45 |
Повторность |
14,7*** |
18*** |
10*** |
1,2 |
24,9*** |
- |
2,1 |
12,7*** |
12,0*** |
- |
24,4*** |
2 |
0,27 |
12,3*** |
12,1*** |
6,89*** |
Остаточная |
82,4 |
76,7 |
79,7 |
97,03 |
69,4 |
76,6 |
97,9 |
80,5 |
88,0 |
100 |
75,1 |
98 |
98,31 |
87,7 |
85,19 |
91,94 |
Примечание - * p<0,05, ** p<0,01, *** p<0,001. |
Библиографический список
- Агроклиматический справочник по Свердловской области. – Л.: Гидрометеоиздат, 1962. – 196 с.
- Бондаренко, А.С. Проблемы реализации “индивидуальной” и “популяционной” схем селекции древесных пород / А.С. Бондаренко // Леса Евразии в XXI веке: Восток-Запад. Материалы II Междунар. конфер. молодых ученых, посвящ. Проф. И.К. Пачоскому (1-5 октября 2002 г.) – М.: МГУЛ., 2002. – С.
- Глотов, Н.В. Естественноисторическое и популяционное исследование скального дуба (Quercus petraea Liebl.) на Северно-Западном Кавказе / Н.В. Глотов, Л.Ф. Семериков, А.В. Верещагин // Журн. общ. биологии, 1975. – Т.36. - №4. – С. 537-557.
- Духарев, В.А. Естественный мутационный процесс в популяциях сосны обыкновенной / В.А.Духарев, Л.Ф. Правдин // Лесоведение, 1983. – №4. – С. 10-14.
- Колесников, Б.П. Леса Свердловской области: Леса СССР / Б.П. Колесников – М.: Наука. – Т.4. - 1969. – С. 64-124.
- Лебедев, Ю.В. Создание постоянной лесосеменной базы в Свердловской области / Ю.В. Лебедев, Т.И. Заровнятных // Лесное селекционное семеноводство: опыт и перспективы. – Екатеринбург: Полиграфист, 2000. – С. 16-18.
- Мамаев, С.А. Закономерности внутривидовой изменчивости семейства Pinaceae на Урале: автореф. дис. … д-ра биол. наук. / С.А. Мамаев Свердловск, 1970. – 54 с.
- Мамаев, С.А. Формы внутривидовой изменчивости древесных растений (на примере семейства Pinacea на Урале) [текст] / С.А. Мамаев. – М.: Наука, 1972. – 283 с.
- Махнев, А.К. Внутривидовая изменчивость и популяционная структура берез секции Albae и Nanae [текст] / А.К. Махнев. – М.: Наука, 1987. – 128 с.
- Основные положения методики закладки испытательных культур плюсовых деревьев главных лесообразующих пород. – 1979. – 19 с.
- Правдин, Л.Ф. Внутривидовая систематика и ее значение для селекции / Л.Ф. Правдин // Проблемы современной ботаники. – М.; Л.: Наука, 1965. – Т.1. – С. 73-76.
- Правдин, Л.Ф. Сосна обыкновенная: Изменчивость, внутривидовая систематика и селекция [текст] / Л.Ф. Правдин. – М.: Наука, 1964. – 191 с.
- Райт, Дж. В. Введение в лесную генетику. Перевод с английского [текст] / Дж. В. Райт.– М., 1978. – 470 с.
- Рокицкий, П.Ф. Биологическая статистика [текст] / П.Ф. Рокицкий. Изд. 3-е, испр. – Минск: «Вышэйш. Школа», 1973. – 320 с.
- Ромедер, Э. Генетика и селекция лесных пород. (Перевод с немец. С.М. Запалова, Х.Я. Бронзовой) [текст] / Э. Ромедер, Г. Шёнбах. – М., 1962. – 268 с.
- Санников, С.Н. Дифференциация популяций сосны обыкновенной [текст] / С.Н. Санников, И.В. Петрова. - Екатеринбург: УрО РАН, 2003. - 247 с.
- Семериков, Л.Ф. Популяционная структура древесных растений (на примере видов дуба европейской части СССР и Кавказа) [текст] / Л.Ф. Семериков. – М.: Наука, 1986. – 163 с.
- Скворцов, А.К. Внутривидовые единицы в ботанической систематике / А.К. Скворцов // Тр. Ин-та экологии растений и животных. - УНЦ АН СССР. 1974. - Вып. 90. С. 46-50.
- Шварц, С.С. Экологические закономерности эволюции [текст] / С.С. Шварц .– М.: Наука, 1980. – 277 с.
- Янбаев, Ю.А. Эколого-популяционные аспекты адаптации лесообразующих видов к условиям природной и техногенной среды: автореф. дис. … д-ра биол. наук./ Ю.А Янбаев. - Тольятти, 2002. – 35 с.
|


