|
"Хвойные бореальной зоны" 2007г.,№2-3, с. 174-178
ОЦЕНКА РОСТА ПОЛУСИБСОВОГО ПОТОМСТВА СОСНЫ КЕДРОВОЙ СИБИРСКОЙ В ОТКРЫТОМ ГРУНТЕ И ТЕПЛИЦЕ
ДанченкоА.М, Кабанова С.А.*
Томский государственный университет, Томск, Россия
*РПГ Научно-производственного центра лесного хозяйства
634050, Томск, пр. Ленина, 36, т. 529 444, факс 529 853, mtd2005@sibmail.com
Проведена сравнительная оценка роста полусибсового потомства 63 семей в открытом грунте и теплице. Установлено, что изменчивость сеянцев с возрастом уменьшается, при этом наибольший размах наблюдается у растений, выращенных в теплице. В основном же уровень изменчивости высоты полусибсовых растений низкий. Очень низким и низким уровнем изменчивости обладали материнские деревья по диаметру шишек, числу в ней чешуй и массы полнозернистых семян. Асимметрия рядов распределения у большинства признаков носит умеренный характер. Ранг среднего у большинства признаков находится в пределах 46–54%. В результате трехлетнего испытания выявлено 12 наиболее перспективных из них, обладающих большой пластичностью, широкой нормой реакции и возрастной стабильностью темпа прироста. Экспериментальные данные свидетельствуют о возрастной стабильности средних значений признаков роста у отдельных семей, что позволяет в 3-летнем возрасте выявить определенные различия в реакции семей на различные фоны и проводить первичный отбор на универсальную приспособленность по признаку потенциальной быстроты роста. Исключение потомств с пониженными показателями темпа прироста обеспечит снижение затрат на дальнейшее их испытание в культурах. Учитывая специфику сосны кедровой сибирской, закладку испытательных культур следует проводить по двум целевым назначениям – на рост и орехопродуктивность, используя в роли регулятора при этом густоту расположения растений на площади.
The growth oi semisib posterity from 63 families was measured in a greenhouse and open ground and the results obtained were compared. The triennial observations revealed 12 most promising generation;) which exhibited high adaptability, expensive reaction norm and high age stability of increment rate. It is suggested that a primary selection be made to identify universally adaptable posterity at an early age. In addition, the experimental plantation from the selected families should be formed to reveal their rapid growth and high seed productivity.
В настоящее время лесная селекция базируется на подборе и выращивании на лесосеменных плантациях, предварительно не проверенных по потомству клонов плюсовых деревьев. Имеется в виду, что такая проверка должна осуществляться после создания плантаций. На основании результатов проверки отбираются элитные деревья и создаются семенные плантации второго поколения.
На различных селекционных объектах выявлено, что качество и продуктивность потомств при семенном размножении растений зависит от комбинационной способности родителей (Турбин, Хотылева, 1961; Березин, 1981; Данченко, 1981). Хотя все признаки имеют генетическую природу, конкретное их проявление при половом размножении зависит от характера взаимодействия генов и условий, в которых растет потомство. Так как древесные размножаются путем перекрестного опыления, а при этом происходит рекомбинация генов, сохранить признаки отобранных форм и материнских деревьев в неизменном виде далеко не всегда представляется возможным. Вероятно, наиболее перспективен не простой отбор плюсовых деревьев, а подбор родителей на комбинационную способность и отбор быстрорастущих сеянцев, выращенных на выровненном агрофоне с последующим их размножением вегетативным путем.
Учитывая сказанное выше, цель настоящей работы заключалась в выявлении возрастной стабильности признаков роста потомства в разных условиях среды для ускоренной оценки их потенциальной продуктивности. Исследования были направлены на поиск генетических ресурсов на разработку программы ускоренного получения эффекта на плантациях целевого назначения.
Объекты и методика
Для выявления характера наследования быстроты роста семенным потомством в качестве моделей отобраны на постоянных пробных площадях материнские деревья, которые имели близкий возраст (80±5 лет) и примерно одинаковые условия конкуренции. При отборе основное внимание обращалось на высоту, диаметр дерева, толщину ветвей, плодоношение, поскольку именно эти параметры являются, с точки зрения хозяйственной ценности, наиболее важными; дополнительные учитывались как возможные диагностические индикаторы. В основном для исследований использовались деревья, контрастные с основным показателям, т.е. принадлежащие к краям распределения по какому-либо показателю, и только часть из взятых моделей относилась к средним и близким к ним по количественным признакам. Перед посевом семена и шишки этих деревьев тщательно изучались с целью создания банка данных и последующего анализа наследуемости основных параметров генеративных органов в испытательных культурах.
Изучению подлежали следующие признаки: масса, шишки, длина, диаметр шишки, расстояние от основания до наиболее широкой части шишки (форма шишки), число чешуи в шишке, число фертильных чешуи, общее число семян в шишке, число полнозернистых семян, масса развитых семян, масса полнозернистых семян, масса одного полнозернистото семени (Данченко, Арцимович, 1990). С целью изучения особенностей реакции, потомства всех семей выращивались одновременно – в условиях открытого грунта и теплицы. От каждого дерева семена высевались блоками точечным методом с размещением растений на расстоянии 5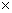 5 см. Повторность размещения рендомизирована по схеме трехкомплексной решетки. От каждого дерева выращивали не менее 64–100 растений. В процессе роста сеянцев ежегодно снималась серия измерений. Обработка экспериментального материала и сравнительный анализ испытуемых семей проводились по стандартной программе. Для оценки взаимосвязи признаков применяли вычисленные парные и ранговые коэффициенты корреляции (r). Оценку уровней изменчивости (Cv) выполняли по шкале С. А. Мамаева (1972). При анализе рядов распределения для измерения степени асимметрии (Ас) использовали нормированный момент третьего порядка, при характеристике крутизны ряда (Е) – нормированный момент четвертого порядка. Для выявления взаимосвязей признаков и поиска информативного из них применяли метод главных компонент. При этом рассмотрению подлежали лишь те компоненты, дисперсия которых составляла более 80% общей. Отбор перспективных семей осуществлялся по индексной оценке темпа прироста в различном возрасте (Роне, Кавац, Бауманис, 1976). 5 см. Повторность размещения рендомизирована по схеме трехкомплексной решетки. От каждого дерева выращивали не менее 64–100 растений. В процессе роста сеянцев ежегодно снималась серия измерений. Обработка экспериментального материала и сравнительный анализ испытуемых семей проводились по стандартной программе. Для оценки взаимосвязи признаков применяли вычисленные парные и ранговые коэффициенты корреляции (r). Оценку уровней изменчивости (Cv) выполняли по шкале С. А. Мамаева (1972). При анализе рядов распределения для измерения степени асимметрии (Ас) использовали нормированный момент третьего порядка, при характеристике крутизны ряда (Е) – нормированный момент четвертого порядка. Для выявления взаимосвязей признаков и поиска информативного из них применяли метод главных компонент. При этом рассмотрению подлежали лишь те компоненты, дисперсия которых составляла более 80% общей. Отбор перспективных семей осуществлялся по индексной оценке темпа прироста в различном возрасте (Роне, Кавац, Бауманис, 1976).
Результаты и обсуждение
Проблема ускоренной оценки селекционного материала является одной из актуальных в настоящее время. Оценка генетических параметров отобранных деревьев проводится как без смены поколений (Драгавцев, 1972; Сахаров, 1974; Петров, 1978), так и по потомства. Разработана и на основе оценки потомств ели, сосны, березы, дугласии апробирована индексная оценка потомств (Роне, Кавац, Бауманис, 1976; Данченко, Дубынин, 1986). Многочисленными исследованиями (Исаков, 1971; Исаков, Пугач, 1974; Долголиков, 1974; Бауманис и др., 1975; Давыдова, 1977, 1980; Молотков, Грицайчук, 1977; .Мухаев, 1978; Этверк, 1974; Василевская, 1983; Шеверножук и др., 1987; Матвеева, 1990 и др.) установлено, что быстрота роста, форма ствола и кроны, толщина сучьев и другие морфологические признаки находятся под определенным генетическим контролем. Экологическое испытание полусибсов сосны кедровой сибирской в питомнике первичного отбора путем регистрации ростовых реакций позволяет в 3-летнем возрасте выявить определенные различия в норме реакции отдельных семей на различные фоны. На основании экспериментального материала установлено, что изменчивость растений с возрастом уменьшается, при этом наибольший размах наблюдается у растений, выращенных в тепличных условиях. В основном же уровень изменчивости высоты растений низкий (таблица 1). В распределении растений по высоте во всех случаях наблюдается лево- и правосторонняя асимметрия, хотя в целом вариационные ряды распределения можно считать симметричными (Ас/ Ас<3) с доверительной вероятностью 0,99. По крутизне рядов распределения выделяются островершинностью двухлетние сеянцы в открытом грунте, а плосковершинностью в тепличных условиях. Коэффициент эксцесса, как правило, при больших значениях третьего основного момента, возрастает. Ранг среднего колеблется в пределах 50% и только у двухлетних растений он несколько выше. В рядах с отрицательной асимметрией величина ранга менее 50%. Таким образом, при равномерной густоте произрастания растений, независимо от экологических условий их выращивания, ряды распределения по высоте близки к нормальным. Анализ индивидуальной изменчивости признаков шишек и семян материнских деревьев показал, что превышение максимального значения признака над его минимальным показателем достигает 4 раз по массе развитых семян в шишке и по массе полнозернистых семян. Ас<3) с доверительной вероятностью 0,99. По крутизне рядов распределения выделяются островершинностью двухлетние сеянцы в открытом грунте, а плосковершинностью в тепличных условиях. Коэффициент эксцесса, как правило, при больших значениях третьего основного момента, возрастает. Ранг среднего колеблется в пределах 50% и только у двухлетних растений он несколько выше. В рядах с отрицательной асимметрией величина ранга менее 50%. Таким образом, при равномерной густоте произрастания растений, независимо от экологических условий их выращивания, ряды распределения по высоте близки к нормальным. Анализ индивидуальной изменчивости признаков шишек и семян материнских деревьев показал, что превышение максимального значения признака над его минимальным показателем достигает 4 раз по массе развитых семян в шишке и по массе полнозернистых семян.
Превышение более чем в 2 раза характерно для числа полнозернистых семян в шишке, общего числа семян в шишке и массы шишки. В меньшей степени проявляется разнообразие материнских деревьев по числу семенных чешуи в шишке, диаметру, длине и форме шишки. Изменчивость признаков характеризуется следующими показателями. Очень низким разнообразием (Cv=5,8%) обладают особи по диаметру шишки. Низкий уровень изменчивости у признаков длины шишки, числа в ней чешуи и массы полнозернистого семени
Средний уровень изменчивости присущ остальным анализируемым признакам. Таким образом, индивидуальная изменчивость генеративных органов не превышает средний уровень. Характеризуя ряды распределения у признаков, констатируем, что асимметрия рядов носит умеренный характер у большинства признаков с правосторонней и только у формы шишек, числа полнозернистых семян и их массы – с левосторонней асимметрией. Плосковершинность наблюдается у формы шишек, числа чешуи и массы семян. Ранг среднего у большинства признаков находится в пределах 50,3–54,2%, у формы шишек, числа и массы полнозернистых семян он 45,6–48,9%.Входящие в выборку материнские особи образуют довольно гетерогенную совокупность, внутри которой они имеют достоверные различия между собой по изучаемым признакам (Fфакт.=4,8–12,5; F01=1,57). Сила влияния организованных в опыте факторов у большинства признаков средняя (0,44–0,57).
Сравнительный анализ изменчивости признаков по годам наблюдений показал, что у изучаемых деревьев в течение двух лет не отмечалось существенных различий (Fфакт.=0,168–2,5;F05=4,2) по форме шишки, числу чешуи в шишке и массе одного полнозернистого семени.
Таблица 1 – Ряды распределения высоты полусибсового потомства в открытом грунте и теплице
Возраст, лет |
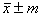
|
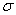
|
Сv |
P |
As |
E |
R |
Открытый грунт |
1 |
3,2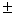 0,1 – 4,5 0,1 – 4,5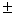 0,1 0,1 |
0,52 – 0,82 |
13,9 –21,6 |
1,3 – 2,2 |
– 0,05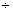 0,86 0,86 |
– 1,15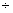 0,62 0,62 |
49,7 – 54,5 |
2 |
5,7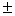 0,1 – 9,2 0,1 – 9,2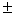 0,1 0,1 |
0,68 – 1,84 |
9,1 – 22,7 |
0,9 –2,2 |
– 0,57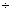 0,90 0,90 |
– 1,23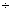 2,91 2,91 |
49,3 – 56,0 |
3 |
10,3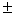 0,2 – 18,1 0,2 – 18,1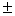 0,2 0,2 |
1,22 – 1,92 |
8,5 –18,6 |
0,8 –1,8 |
– 0,42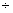 0,26 0,26 |
– 1,03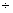 0,87 0,87 |
47,2 – 51,7 |
Теплица |
1 |
3,3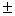 0,1 – 4,0 0,1 – 4,0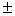 0,1 0,1 |
0,43 – 0,69 |
12,2 – 18,9 |
1,2 – 1,9 |
– 0,14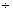 0,47 0,47 |
– 0,91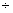 0,14 0,14 |
49,0 – 53,1 |
2 |
5,7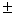 0,1 – 6,7 0,1 – 6,7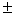 0,1 0,1 |
0,66 – 0,79 |
9,8 – 12,6 |
0,9 – 1,2 |
– 0,14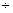 1,77 1,77 |
– 0,45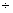 8,11 8,11 |
49,0 – 61,8 |
3 |
11,3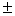 0,1 – 14,0 0,1 – 14,0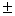 0,2 0,2 |
1,42 – 1,83 |
9,9 – 12,6 |
1,0 – 1,3 |
– 0,17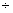 0,57 0,57 |
– 0,76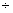 0,73 0,73 |
49,2 – 53,7 |
По данным признакам деревья относительно четко сохраняли свой ранг (rs=0,50–0,58>F05), за исключением числа фертильных чешуи в шишке (rs=0,34). При достоверных различиях диаметра шишки по годам наблюдений (Fфакт.=20,5; F05=42), деревья все же очень четко сохраняют свой ранг в ряду распределения (rs=0,70>r05). Таким образом, условия смежных вегетационных периодов в меньшей степени оказывают влияние на формирование перечисленных признаков, чем их генотипические особенности.
Существенно изменялись масса шишки, длина шишки, число семян и их масса (Fфакт.=7,6–149,6; F05=4,2), при этом неустойчив и ранг деревьев по данным признакам (r=0,03–0,33). Это свидетельствует о различной реакции деревьев на формирующие условия вегетационного периода. В то же время погодные условия значительно отражались на доле фенотипической изменчивости.
На основании экспериментального материала установлено, что в открытом грунте наиболее перспективными по росту в высоту (J1) оказались семьи 167, 121, 174, 49 и 113, занимающие первые пять мест (интенсивность отбора около 10%) из общей совокупности испытуемых 63 семей. Перспективны также семьи 1, 188, 148, 160, 156, 451, 57 при интенсивности отбора в 20%. Однако кроме общих показателей роста сеянцев важно знать и темп их прироста (J2). Установлено, что различия потомств в темпах прироста за период до его кульминации и без смены экологических условий (пересадка) удовлетворительно описываются регрессионным уравнением y=bo+b1x. Высокие значения имеют потомства 167, 121, 49, 174. Высокий темп прироста у потомства 57. Из общей совокупности перспективных семей выпали семьи 160 и 441, которые соответственно стали иметь ранги 14 и 27. В то же время по общей оценке в эту группу вошли семьи 89, 142. Остальные с незначительным изменением рангов сохранились в перспективной по росту группе.
При выращивании сеянцев в теплице группа наиболее перспективных по росту семей несколько изменилась, хотя в ней сохранились семьи 57, 121, 49. Наиболее устойчивыми по росту и темпу прироста в теплице оказались семьи 6, 39, 14, которые обладают на» большей нормой реакции. По росту в высоту в группу перспективных семей входили 157, 139, 16, 38, при общей же оценке они выпали и замещены семьями 2, 424, 121, 17.
Обобщенная оценка полусибсового потомства на двух экологических фонах выявила группу наиболее перспективных семей, в которую вошли 57, 121, 49, 182, 2, 174, 89, 167, 38, 14, 39 и 423 семьи. К особо ценным семьям, обладающим большой пластичностью и широкой нормой реакции, можно отнести 121, 49 и 57, в меньшей мере – 167, 174, 182.
По суммарной комплексной оценке из 28 перспективных семей в группу особо ценных вошли 6 семей при отборе в открытом грунте (57, 121, 49, 174, 188, 167) и 5 из теплицы (182, 6, 49, 14, 39), или около 35%. В целом же из всех 63 испытанных семей это составляет 16%. Проявили нестабильность семьи: 113, 1, 189, 148, 160, 156, 441 и 142, выращенные в открытом грунте, и 2, 22, 157, 139, 16, 37, 424, 17, 423 – из теплицы. Следовательно, взаимодействие потомства с условиями выращивания выявляет оптимальные экологические нормы для роста испытанного потомства.
Таким образом, при последующей закладке культур в испытание следует включить весь перечень перспективных потомств (28 из таблицы 2) или, при недостатке площадей, следует продолжить испытание только выявленных по обобщенной оценке (57, 121, 49, 182, 2, 174, 89, 167, 38, 14, 39,423). При закладке испытательных культур следует предусмотреть два варианта: на быстроту роста со схемой закладки 3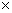 3 м (1100 шт./га) и плодоношение – при размещении 6 3 м (1100 шт./га) и плодоношение – при размещении 6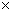 6 или 8 6 или 8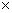 8 м. 8 м.
Таблица 2 – Комплексная оценка перспективных семей
Условия выращивания |
Ранг семьи |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
Открытый грунт, J1 |
167 |
121 |
174 |
49 |
113 |
1 |
188 |
148 |
160 |
156 |
441 |
57 |
Открытый грунт, J1+ J2 |
167 |
121 |
49 |
57 |
174 |
148 |
113 |
188 |
156 |
1 |
89 |
142 |
Теплица, J1 |
6 |
14 |
22 |
39 |
157 |
49 |
139 |
16 |
37 |
38 |
182 |
423 |
Теплица, J1+ J2 |
57 |
6 |
39 |
2 |
14 |
182 |
424 |
121 |
17 |
49 |
22 |
37 |
Обобщенная оценка |
57 |
121 |
49 |
182 |
2 |
174 |
89 |
167 |
38 |
14 |
39 |
423 |
Таблица 3– Корреляционная связь между признаками материнских деревьев и ростом
потомства
Признак |
Номер признака |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
1 |
Масса шишки |
– |
|
|
|
|
|
|
|
2 |
Длина шишки |
0,686 |
– |
|
|
|
|
|
|
3 |
Число семян в шишке |
0,744 |
0,611 |
– |
|
|
|
|
|
4 |
Число полнозернистых семян в шишке |
0,786 |
0,497 |
0,809 |
– |
|
|
|
|
5 |
Масса полнозернистых семян |
0,867 |
0,526 |
0,779 |
0,965 |
– |
|
|
|
Высота потомства: |
6 |
1-летнего |
-0,146 |
-0,057 |
-0,113 |
-0,142 |
-0,111 |
– |
|
|
7 |
2-летнего |
-0,356 |
-0,333 |
-0,330 |
-0,262 |
-0,271 |
0,512 |
– |
|
8 |
3-летнего |
-0,156 |
-0,207 |
-0,346 |
-0,135 |
-0,126 |
0,125 |
0,580 |
– |
Таблица 4 – Результаты анализа методом главных компонент связей признаков
Признаки |
Главные компоненты |
1 |
2 |
3 |
Масса шишки |
0,953 |
- 0,111 |
0,031 |
Длина шишки |
0,748 |
- 0,041 |
- 0,092 |
Число семян в шишке |
0,915 |
0,062 |
- 0,061 |
Число полнозернистых семян в шишке |
0,920 |
0,162 |
0,064 |
Масса полнозернистых семян |
0,953 |
0,0,176 |
0,058 |
Высота потомства: |
1-летнего |
- 0,259 |
0,393 |
0,233 |
2-летнего |
- 0,534 |
0,474 |
0,001 |
3-летнего |
- 0,372 |
0,405 |
0,282 |
Доля влияния компоненты, % |
52,91 |
19,7 |
11,5 |
В таблице 3 приведены данные изучения фенотипических связей между признаками генеративных органов материнских деревьев и ростом потомства в первые три года. На основе корреляционной матрицы проведены расчеты главных компонент. В таблице 4 представлены весовые коэффициенты, оценивающие корреляцию между исходными признаками и выделенными компонентами. Главные компоненты рассчитывались для всех исходных признаков, однако, как видно из таблица 2, первые три охватывают более 80% всей изменчивости. Наибольший вес имеет первая главная компонента, доля влияния которой составляет 52,9% общей изменчивости и которую можно интерпретировать как компоненту массы шишки. В эту компоненту с большими весовыми коэффициентами входят массовые и числовые показатели семян. Рост сеянцев имеет аллометрическую связь с признаками материнских деревьев, что особенно проявляется в двухлетнем возрасте. Вторая главная компонента характеризуется ростовыми показателями потомства, ведущая роль при этом принадлежит высоте двухлетних растений. С высокими весовыми показателями в эту компоненту входят масса полнозернистых семян и их число в шишке. И, наконец, третью компоненту можно назвать компонентой высоты трехлетних сеянцев. Таким образом, можно видеть, что на ранних стадиях развития потомства в определенной степени на его рост оказывают влияние некоторые характеристики признаков семян. Очевидно, двухлетняя высота растений является определяющей в общей трехлетней оценке потомства, но данный вывод следует проверить на потомствах нескольких генераций.
Заключение
Экспериментальные данные свидетельствуют о возрастной стабильности средних значений признаков роста у отдельных семей, что позволяет в 3-летнем возрасте выявить определенные различия в реакции семей на различные фоны и проводить первичный отбор на универсальную приспособленность по признаку потенциальной быстроты роста. Исключение потомств с пониженными показателями темпа прироста обеспечит снижение затрат на дальнейшее их испытание в культурах.
Учитывая специфику сосны кедровой сибирской, закладку испытательных культур следует проводить по двум целевым назначениям – на рост и орехопродуктивность, используя в роли регулятора густоту расположения растений на площади.
Библиографический список
- Бауманис, И.И. Изменчивость признаков сосны обыкновенной и методика оценки потомства / И.И. Бауманис, М.Г. Паэгле, В.И. Роне // Генетические исследования древесных в Латвийской ССР. – Рига: Знание, 1975. – С. 25–34.
- Березин, Э. Л. Комбинационная способность при внутривидовых скрещиваниях и ее значение для селекции ели / Э. Л. Березин // Разработка систем селекции древесных пород. – Рига, 1981. – Ч. 2. – С. 3–6.
- Василевская, Л. С. Проверка плюсовых деревьев дуба черешчатого по потомству / Л.С. Василевская // Всесоюз. совещ. по лесной генетике, селекции и семеноводству. – Петрозаводск, 1983. – Ч. 2. – С. 44–45.
- Давыдова // Лесоводство и агролесомелиорация. – Киев: Урожай, 1977. – Вып. 48. – С. 10–14.
- Давыдова, И. И. Исследование потомства плюсовых деревьев дуба / И.И. Давыдова // Селекция, генетика и семеноводство как основа создания высокопродуктивных лесов. – М., 1980. – Ч. 2. – С. 101–104.
- Данченко, А. М. Оценка комбинационной способности при внутривидовых скрещиваниях березы повислой / А. М. Данченко // Разработка основ систем селекции древесных пород. – Рига, 1981. – Ч. 2. – С. 10–14.
- Данченко, А.М. Внутрпопуляционная изменчивость характеристик шишек кедра в подзоне южной тайги / А. М. Данченко, Н. Ф.Арцимович // Проблемы кедра: Семеношение и размножение. – Томск, 1990. – Вып. 4. – С. 34–57.
- Данченко, А. М. Рост потомства березы в испытательных культурах Северного Казахстана / А.М. Данченко, Г.Б. Дубынин // Лесовосстановление в Казахстане. – Алма-Ата: Кайнар, 1986. – С. 51–63.
- Долголиков, В. И. О ранней диагностике быстроты роста в высоту у сосны и ели по прямому признаку / В.И. Долголиков // Состояние и перспективы развития лесной генетики, селекции, семеноводства и интродукции. – Рига, 1974. – С. 45–48.
- Драгавцев, В.А. Экспериментальное сопоставление трех принципов оценки генотипической изменчивости количественных признаков в растительных популяциях/ В. А. Драгавцев // Генетика. – 1972. – Т. 8, № 5. – С. 28–34.
- Исаков Ю.Н. Изменчивость двухлетних сеянцев сосны по высоте в зависимости от особенностей материнских деревьев / Ю.Н. Исаков // Пути совершенствования лесосеменного дела в лесхозах лесной зоны. – Киров, 1971. – С. 42–44.
- Исаков, Ю.Н. К использованию коэффициента повторяемости для раннего отбора на быстроту роста у сосны обыкновенной / Ю.Н. Исаков, Г.В Пугач // Состояние и перспективы развития лесной генетики. – Рига, 1974. – С. 61–64.
- Мамаев, С.Л. Формы внутривидовой изменчивости древесных растений (на примере Pinaceaeна Урале) [текст] / С.Л. Мамаев.- М.: Наука, 1972. – 283 с.
- Матвеева, Р. Н. Программа и результаты исследований по выращиванию сортового посадочного материала сосны кедровой сибирской / Р.Н. Матвеева // Проблемы кедра: Региональные программы. – Томск, 1990. – С. 86–93.
- Молотков, П.И. Исследование семенного потомства плюсовых деревьев в Тростянецком лесхоззаге / П.И. Молотков // Лесоводство и агролесомелиорация. – Киев:Урожай, 1977. – Вып. 48. – С. 15–18.
- Мухаев, Б.А. Наследование некоторых материнских признаков семенным потомством берез / Б.А. Мухаев // Бюл. ВНИАЛМИ. – 1978. – Вып. 1. – С. 16–18.
- Петров, С.А. Принципы генетической оценки плюсовых деревьев / С. А. Петров // Лесное хоз-во. – 1978. – № 1. – С. 75–77.
- Роне, В. М. Селекционная оценка потомств сосны и ели / В.М. Роне, Я.Э. Кавац, И.И. Бауманис // Лесоведение. – 1976. – № 5. – С. 30–38.
- Сахаров, В.И. Возможности метода фоновых признаков при изучении феногенетической структуры популяций / В.И. Сахаров // Состояние и перспективы развития лесной генетики, селекции, семеноводства и интродукции: Методы селекции древесных пород. – Рига, 1974. – С. 103–106.
- Турбин, Н.В. О принципах и методах селекции растений на комбинационную способность / Н.В. Турбин, Л.В. Хотылева // Гетерозис: Теория и методы практического использования. – Минск, 1961. – С. 59–110.
- Шеверножук, Р.Г. Испытание полусибсов сосны в фоновом питомнике / Р.Г. Шеверножук, Ю.А. Ломовских, А.А Хиров // Селекция и семеноводство хвойных. – Воронеж, 1987. – С. 25–33.
- Этверк, И. Э. Об оценке производительности плюсовых деревьев ели обыкновенной / И. Э. Этверк // Наука – производству. – Каунас, 1974. – Вып. 2. – С. 25–30.
|


