|
Биологическая продуктивность лесных культур на бореальном экотоне
Усольцев В.А.1,2, Терехов Г.Г.2, Ненашев Н.С.3, Пальмова Н.В.4, Балицкий М.И.4,. Касаткин А.С4, Лысенко Д.И.1, Канунникова О.В.1, Кузьмин Н.И.5
1 Уральский государственный лесотехнический университет, Екатеринбург, Россия
2 Ботанический сад Уральского отделения РАН, Екатеринбург, Россия
3 Омский государственный агроуниверситет, Омск, Россия
4 Оренбургский государственный агроуниверситет, Оренбург, Россия
5 Агентство лесного хозяйства по Оренбургской области, Оренбург, Россия
Дана характеристика иерархической системы трех экотонов – зонального, подзонального и фитоценотического. Исследована биопродуктивность культур сосны обыкновенной на подзональном экотоне Урало-Тургайско-Западносибирского региона. При отсутствии различий продуктивности культур высших (Ia-I) классов бонитета в пределах трех таксонов (принцип взаимозаменяемости экологических факторов), продуктивность модальных древостоев культур закономерно снижается в направлении с севера на юг. Охарактеризованы зоны перекрытия петли гистерезиса (триггерной полосы) на экотоне в терминах фитомассы и ее годичной продукции.
Hierarchical system of three ecotones – zonal, sub-zonal, and phytocoenotic – has been given. Biological productivity of Pinus sylvestris plantations on the sub-zonal ecotone of the Ural-Turgai-West-Siberian region has been studied. When among three taxons of the sub-zonal ecotone any difference in productivity of plantations of highest (Ia-I) site indices has not been revealed, modal stand productivity is decreased in direction from the north to the south. The overlapping ranges of the ecotone trigger belt in terms of plantation biomass and NPP have been shown.
Введение
Неизбежность глобального потепления, обусловленного ростом концентрации атмосферного СО2 из-за сжигания все возрастающего количества ископаемого углеродного топлива была показана еще в конце XIX в. в работах одного из основоположников теории неравновесной кинетики Сванте Аррениуса (цит. по: Будыко, 1974), которые в дальнейшем были забыты. Прогнозируемое сегодня климатическое потепление, довольно резкое в историческом масштабе, может вызвать радикальные изменения в растительном покрове (Уткин, 2001).
Для оценки климатогенных изменений в структуре лесного покрова особый интерес представляют системы экотонов (переходов между сообществами растений на высотных и зональных границах бореальных лесов) поскольку именно здесь наблюдается наиболее выраженная реакция растительности на изменение климата. Поэтому изучению пространственно-временной динамики переходных растительных сообществ уделяется исключительно большое внимание (Korner, 1998; Bugmann, Pfister, 2000; Шиятов и др., 2005). Эта динамика должна оцениваться в терминах биологической продуктивности, поскольку биопродуктивность, во-первых, является определяющей характеристикой растительного покрова и, во-вторых, ее показатели менее всего изучены именно на зональных и верхних пределах лесного ареала, что вносит неопределенность в оценку продукционного потенциала покрытых лесом площадей.
Представим в некотором приближении переходные состояния на южном пределе лесов Северной Евразии в виде своеобразной иерархической системы пространственных экотонов:
1) зональный экотон лес - степь как переходное состояние растительных формаций между таежной и степной природными зонами;
2) подзональный (бореальный) экотон - как переходное состояние лесных формаций между лесными подзонами (в европейской России – это экотоны южная тайга - широколиственные леса или широколиственные леса - лесостепь; в Сибири – это предлесостепь как переходная полоса между южной тайгой и лесостепью);
3) фитоценотический экотон - опушка как переходное состояние между лесными и степными (луговыми) фитоценозами.
Зональный экотон. Первые и довольно противоречивые описания переходных состояний растительности на южном пределе бореальных лесов, или на зональном экотоне, появились в конце XIX столетия (Пермский, 1876; Коржинский, 1888, 1891; Танфильев, 1894; Богатов, 1899). Дебаты шли по двум основным вопросам: а) каковы причины безлесия степей и б) была ли степь извечно безлесной? Или, в современных терминах переходных состояний, каковы причины возникновения и существования экотона лес-степь, и существовал ли он всегда или появился в историческое время под влиянием климатических изменений либо антропогенных воздействий?
Мнения ученых разделились: одни доказывали, что леса существовали на степном юге России 700-1000 лет назад (П. Голубовский: цит. по: Собичевский, 1890) и что нынешнее безлесие степей – явление искусственное, вызванное вмешательством человека (Палимпсестов, 1882), а другие утверждали, что южная Россия во все исторические времена была безлесной степью (Bar, 1856).
Рассматривалось несколько причин безлесия степей.
Климат. Сторонники климатической теории безлесия степей (Bar, 1856; А. Гризебах (цит. по: Танфильев, 1894); Г.Е. Щуровский (цит. по: Собичевский, 1890) считали основным фактором дефицит атмосферной и почвенной влаги, а А. Миддендорф (Middendorf, 1864) – сильные ветры, которые сдувают со степи снег, являясь таким образом и следствием, и причиной существования степей. Но это противоречило выводу А. Богатова (1899), что «степная и лесная формации для своего существования требуют одинаковых условий местообитания» (с. 560).
Почвы. Ф. Рупрехт (1866) и А. Майков (1874) видели главную причину безлесия в свойствах чернозема, Ф. Тицман (Teezman, 1845) – в непроницаемости подпочвы степей, а П.А.Костычев (1886) - в различии механического состава степных и лесных почв. Последний показал сходство по климатическим показателям черноземных и лесных регионов и утверждал, что степные и лесные формации могут существовать при однородных климатических условиях. Но это расходится с мнением сторонников климатической теории.
По Г.И. Танфильеву (1894), причина безлесия степей – в специфике химического состава степных почв, а именно, в наличии карбонатов, а по В.В. Докучаеву (1889) – в «известной солености почв». Но А. Богатов (1899) приводит факты успешного произрастания лесов не только на почвах с высоким содержанием карбонатов, но и на солонцоватых, и полагает, что «выщелоченность лесных почв есть следствие, но не причина существования на них лесов» (с. 570).
Топография местности. Согласно наблюдениям А. Богатова (1899), лесные острова в степи повсеместно совпадают с областями наибольшего развития оврагов и балок, и местности, наиболее овражистые, оказываются и наиболее лесистыми. Он пришел к выводу, что «не только распределение и расположение лесов, но и состав их находится в зависимости от топографии местности» (с. 559-560). Аналогичного мнения придерживался А.Н. Краснов (1890).
Конкурентная борьба между растительными формациями. Наиболее последовательным сторонником подобной версии был С.И. Коржинский (1891): «…В нашей полосе распределение лесных и степных формаций не зависит непосредственно ни от климата, ни от топографического характера местности, ни от природы и свойств субстрата, но только от условий и хода взаимной борьбы за существование» (с. 172 и 173). Конкурентная борьба между лесной и степной растительностью отмечалась также в Сибири (Фрейдин, 1900). Однако Г.И. Танфильев (1894) отрицал такое толкование и комментировал его следующим образом: «Объяснять данное распределение растительных формаций отвлеченной борьбой за существование равносильно отказу от какого бы то ни было объяснения» (с. 246).
Обсудив «наиболее выдающиеся теории», объясняющие «загадочные особенности степей», А. Богатов (1899) не соглашается ни с одной из них. Его концепция остается актуальной по сей день: «Такие сложные явления, как рассмотренные выше особенности степей, вряд ли можно объяснить одной какой-либо причиной. Сложное явление всегда есть результат совокупного действия многих причин. Общий недостаток всех рассмотренных теорий заключается в том, что авторы их, увлекшись одной какой-либо причиной, старались все остальные держать на приличном отдалении. Очень может быть, что причиною этих явлений следует считать и климат, и почву, и рельеф местности. Климат, конечно, влияет на географическое распространение растений по поверхности Земли; в пределах же климатических поясов распределение растений зависит от почвы, а также от истории и топографии страны» (с. 575).
Е.М. Лавренко (1940) по существу подтверждает общую концепцию А. Богатова на примере экотона лес-степь. Не отрицая роли климата к его формировании, Е.М. Лавренко подразделяет абиотические факторы, определяющие взаимодействие леса и степи, на два уровня: «…В ряде районов лесостепной полосы участки леса и степи (в настоящий момент, вернее, лесных и степных почв) сменяют друг друга вне зависимости от условий рельефа и характера подпочв, в то время как в пределах южных степей леса всегда связаны с определенными условиями рельефа (балки, долины рек) или подпочвы (более легкие или каменистые почвы)» (с. 16).
Подзональный (бореальный) экотон. В последние годы в биогеографии получило распространение понятие бореальный экотон как векторная природно-территориальная составляющая поясного ранга, простирающаяся полосой от Прибалтики до Предбайкалья (Коломыц и др., 1993). Ординация подзональных типов лесного покрова Европейской России, выполненная Н.И. Базилевич с соавторами (1986), выявила их существенное взаимное перекрытие в полосе бореального экотона, а следовательно и неизбежную конкуренцию между видами и сообществами древесных растений (Сукачев, 1972). По Э.Г. Коломыцу с соавторами (1993), бореальный экотон – это «не просто переходная полоса между двумя относительно однородными биотическими сообществами. Это вполне самостоятельное структурное образование биосферы, нередко отличающееся от соседних однородных экосистем повышенной сложностью ценотической структуры и более интенсивным вещественно-энергетическим обменом, а в конечном итоге и более высокой биопродуктивностью» (с. 11).
При описании взаимодействий лесных экосистем на бореальном экотоне показано (Варлыгин, Базилевич, 1992; Коломыц и др., 1993), что здесь происходит смена ролей тепла и влаги как фоновыхлимитирующих факторов биологической продуктивности. Увеличение биопродуктивности лесного покрова в таежном поясе происходит за счет роста теплообеспеченности, а в лесостепном – при возрастании обеспеченности влагой (Варлыгин, Базилевич, 1992). В таежном поясе рост увлажнения ведет к снижению биопродуктивности, а в лесостепном – напротив, к ее увеличению.
Э.Г. Коломыц с соавторами (1993) на примере юга европейской России, придерживаясь концепций А. Богатова (1899) и Е.М. Лавренко (1940), показали, что действующие по профилю бореальной зоны фоновые лимитирующие факторы (тепло и влага) в пределах бореального экотона модифицируются системой локальных форм рельефа и эдафотопа, и фоновая ландшафтно-климатическая однородность подзонального экотона здесь сочетается с резко выраженными локальными природно-территориальными контрастами и соответственно с мозаичностью ландшафтной структуры экотона.
Исследуя биопродуктивность сосняков Окско-Мокшинского песчаного полесья на подзональном экотоне, т.е. между южной тайгой на севере и широколиственными лесами на юге, Э.Г. Коломыц с соавторами (1993) выявили здесь феномен раздвоения ландшафтно-экологического оптимума, что в общем согласуется как с правилом предварения В.В. Алехина (1935), так и с принципом взаимозаменяемости экологических факторов Л.Г. Раменского (1938): биопродуктивность в условиях плакорного микрорельефа соответствует фоновому показателю южной тайги, а в условиях «неплакорного» микрорельефа – близка к фоновой для широколиственных лесов. В полосе подобных контрастов происходит смена одного фонового ведущего фактора (тепло) другим (влага). В ее локальных местообитаниях недостаточная трофность почвы, как и недостаток атмосферной влагообеспеченности, компенсируются повышенным эдафическим увлажнением. Анализ влияния эдафических и топологических факторов на структурно-функциональную организацию хвойных лесов, показал, что одно и то же фоновое воздействие (например, изменение климата) вызывает неоднозначный локальный отклик: одни экосистемы приближаются к экологическому оптимуму, а другие удаляются от него.
Размах локальных контрастов биологической продуктивности лесных экосистем на бореальном экотоне соизмерим с интервалом продукционного фона от лесотундры до степей. При этом биологическая продуктивность лесного фитоценоза на подзональном экотоне может быть выше, чем на прилегающих к нему подзонах (Коломыц, 1984; Коломыц и др., 1993). Это подтверждается фактическими данными определения фитомассы сосняков, согласно которым наибольшей продуктивностью характеризуются лесные насаждения лесостепи по отношению к лесорастительным таксонам, прилегающим с севера и юга в европейской России (Коломыц и др., 1993) и в Сибири, как Западной (Таран, 1973; Габеев, 1990), так и Средней (Горбатенко, 1975). По Уральскому меридиану наибольшей продуктивностью в лесостепи отличаются также березняки, к северу и югу от которой она закономерно снижается (Усольцев, 2001).
Таким образом, на подзональном (бореальном) экотоне происходит смена ролей тепла и влаги как фоновых лимитирующих факторов, а система локальных форм рельефа и эдафотопа определяет смену ролей локальных лимитирующих факторов – атмосферной влагообеспеченности, гидрологического режима подпочв и трофности почв. По зональному градиенту экологический оптимум для сосны обыкновенной и березы находится в подзоне лесостепи.
Фитоценотический экотон – лесная опушка. Пограничный переход между площадями, занятыми взаимопроникающими на экотон лесными и степными фитоценозами, представляет собой опушку, т.е. узкую переходную зону, приходящуюся на границу раздела типичных степных и типичных лесных почв. Под естественными сосновыми древостоями Аман-Карагайского бора в Тургайском прогибе (51030’ с.ш., 64015’ в.д.) обычны слаборазвитые дерново-боровые почвы с уровнем залегания грунтовых вод до 7-9 м, а на прилегающих остепненных пространствах распространены зональные темно-каштановые почвы, характеризующиеся развитым гумусовымгоризонтом и наличием карбонатных прослоев в подстилающих глинах и суглинках. При благоприятных погодных условиях и высоком уровне грунтовых вод в семенные годы происходит экспансия сосны с дерново-боровых на темно-каштановые почвы. Полоса такого экотона (опушка) довольно узкая, т.е. сомкнутый древостой резко сменяется степным ландшафтом. При средних годовых осадках около 240 мм уровень грунтовых вод в сухие периоды опускается, и сосняки на зональной почве гибнут, уступая место степной растительности, а при благоприятных условиях вновь наступают на степь.
 Полосы названных трех экотонов имеют ширину разных порядков: если зональный и подзональный экотоны простираются соответственно на сотни и десятки километров, то ширина фитоценотического экотона может составлять несколько метров. По градиенту эдафотопа может появиться экотон между жизненными формами, различающимися между собой в меньшей степени, чем лесная и травянистая (кустарниковая) растительность, например, между сосной на песчаном плакоре и березой на пониженных элементах рельефа, ширина которого также может составлять считанные метры. Полосы названных трех экотонов имеют ширину разных порядков: если зональный и подзональный экотоны простираются соответственно на сотни и десятки километров, то ширина фитоценотического экотона может составлять несколько метров. По градиенту эдафотопа может появиться экотон между жизненными формами, различающимися между собой в меньшей степени, чем лесная и травянистая (кустарниковая) растительность, например, между сосной на песчаном плакоре и березой на пониженных элементах рельефа, ширина которого также может составлять считанные метры.
Экотоны всех уровней имеют одну общую особенность – наличие конкурентных отношений между растительными видами и их формациями. Это нашло отражение в концепции критического состояния экосистемы в результате потери устойчивости А.Д. Арманда (1989). Согласно этой концепции, критическим является такое состояние экосистемы, в котором происходит ее качественная перестройка. Пока в экосистеме действует механизм обратной связи, она сохраняет способность к восстановлению и остается в некоторой области устойчивости. В случае разрушения экосистемы и формирования новой структуры имеет место переход в новую область устойчивости. Момент перехода означает критическое состояние, или критическую точку. Такую пару устойчивых экосистем, разделенных критической точкой, называют триггером, или переключателем, что является по существу синонимом петли гистерезиса (нелинейности в среде с памятью) (Ведюшкин, 1992), или «складки» в терминах теории катастроф (Чиллингуорт, 1979).
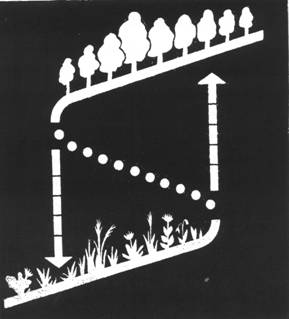
Верхнее и нижнее крылья складки могут соответствовать, например системам лесной фитоценоз и травяной фитоценоз (рисунок 1). Участок оси абсцисс, на котором оба крыла имеют одинаковые координаты, соответствует триггерной полосе. Реальная климатическая граница леса и степи располагается внутри складки. Для определения ширины триггерной полосы на местности необходимо располагать сведениями о возможности восстановления нарушенных ландшафтов в обе стороны от действительной границы леса. При наступании лесной зоны на степь под влиянием изменения климата складка движется вместе с границей и находится в пределах травяных ландшафтов перед фронтом леса, а при наступании степной зоны вся складка находится в пределах леса (Арманд, Кушнарева, 1989).
Представление экотона лес - степь в качестве триггерной полосы, или петли гистерезиса (Арманд, Ведюшкин, 1989; Арманд, Кушнарева, 1989; Ведюшкин, 1992) означает, что его границы по условию пространственного гистерезиса не должны восстанавливаться по прежней траектории после окончания внешнего воздействия, если оно превышает некоторую критическую величину.
Рисунок 1 - Схематическое изображение триггерной системы лес – степь в координатах фитомасса на единице площади – расстояние (Арманд, Кушнарева, 1989). Пояснения см. в тексте
Сказанное можно отнести не только к конкурирующим растительным сообществам, но и к соответствующим эдафическим фонам. Суть процесса на сопряженных категориях почв, объясняемого явлением гистерезиса в «среде с памятью», была изложена А.А. Завалишиным (1936). Критикуя «циклическую» гипотезу о взаимоотношении леса и степи, А.А. Завалишин пишет, что при наступании леса на степь идет процесс деградации (оподзоливания) черноземов, но при обратном переходе - наступании степи на лес процесс реградации почв идет по иной траектории, и чем сильнее оподзолена почва, тем меньше шансов на ее полное восстановление или, иными словами, процесс реградации не может полностью стереть следы деградации.
На основе изложенного выше о переходных состояниях экосистем можно определить экотон как обусловленную действием лимитирующих факторов полосу перехода от доминирования одной жизненной формы к доминированию другой, характеризующуюся наличием конкурентных отношений, от напряженности которых зависит крутизна (резкость) перехода (Арманд, Кушнарева, 1989; Ведюшкин, 1992; Korner, 1998). К разным жизненным формам относят не только деревья и кустарники или деревья и травянистые растения, но и лиственные вечнозеленые и лиственные листопадные или хвойные вечнозеленые и хвойные листопадные (Jobbagy, Jackson, 2000). Имеются виду переходы как в пространстве, так и во времени, причем последние идентифицируются с использованием сравнительно-географического метода А.К. Гумбольдта (Арманд, 1989), или метода пространственных аналогий (Арманд, Кушнарева, 1989).
В литературе упоминаются две взаимоисключающие тенденции в динамике растительности на экотоне в терминах продуктивности: в одних случаях продуктивность сообщества на экотоне выше, чем на прилегающих площадях (Коломыц и др., 1993), а в других – напротив, ниже (Одум, 1975). По-видимому, в случаях, когда конкурируют близкие по морфоструктуре жизненные формы, например, сосна и береза на равнине или разные комбинации кустарниковой растительности в горах Кавказа, то продуктивность сообщества на экотоне выше, чем за его пределами. Это с математически обусловленной неизбежностью следует из хорошо известного феномена: если в пределах некоторого диапазона по оси абсцисс действуют на встречных направлениях два убывающих тренда, то их совокупный эффект дает колоколообразную кривую с максимумом в области пересечения трендов (Кофман, Кузьмичев, 1979).
Если же конкурируют совершенно разные жизненные формы, например, деревья и травы, то имеет место постепенное снижение продуктивности сообщества по направлению от границы леса к границе степной растительности, поскольку продуктивность сообщества на таком экотоне определяется исключительно древесной растительностью, и на ее фоне вклад иной растительности на встречном направлении ничтожно мал.
Таким образом, взаимодействие фитоценозов на экотонах определяется, как и полагал А. Богатов, 1899), и климатом, и почвой, и рельефом местности, но соотношение этих факторов на экотонах разных уровней различное. Конкурентные же взаимодействия между фитоценозами проявляются на экотоне любого уровня.
Наше исследование биологической продуктивности выполнено в пределах подзонального (бореального) экотона в Урало-Тургайско-Западносибирском регионе в лесных культурах сосны обыкновенной на лесопокрытых площадях трех административных областей соответственно в трех лесорастительных таксонах: предлесостепь (Свердловская область, Сухоложский лесхоз), лесостепь (Омская область, Саргатский лесхоз) и сухая степь (Северный Казахстан, Кустанайская область, Семиозерный лесхоз) (рисунок 2). Экотон в данном случае представляет собой переходное состояние между предлесостепью на севере и сухой степью на юге.
Для изучения биопродуктивности лесных культур было, по крайней мере, две причины. Во-первых, биологическая продуктивность лесных культур изучена в целом по стране намного хуже, чем естественных насаждений. Поэтому получение фактических данных по культурам дает возможность заполнить многочисленные «белые пятна» и сопоставить эти данные с аналогичными материалами для естественных насаждений. Во-вторых, лесные культуры, особенно молодые, связывают атмосферный углерод более интенсивно в сравнении с естественными насаждениями. Однако в бореальном экотоне региона углерододепонирующие возможности лесных культур изучены слабо, имеются лишь единичные определения.
Трудности при первых лесопосадках М. Савич (1885) связывает с проявлением «народного невежества»: «Лесоразведение в Войске при своем возникновении не имело за себя симпатий населения; к нему отнеслись, можно сказать, враждебно, но по мере того как благоприятные результаты этого «новшества» делаются очевидными, к нему начинают относиться более благосклонно» (с. 34). Сегодня лесокультурное производство России достигло по Рослесхозу ежегодного объема лесовосстановительных работ на площади около 800 тыс. га, из которых 39 % приходится на сосну обыкновенную (Леса…, 2006).
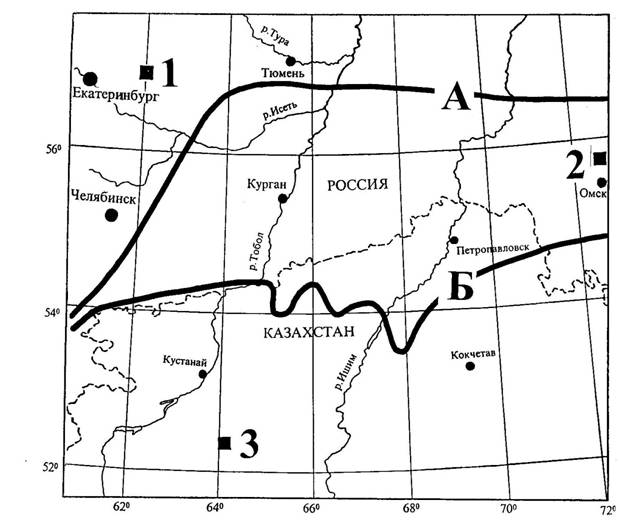
А – южная граница предлесостепи и подтаежных лесов; Б – то же, лесостепи
Рисунок 2 - Расположение мест закладки пробных площадей на бореальном экотоне: 1– Сухоложский лесхоз Свердловской области (предлесостепь), 2 - Саргатский лесхоз Омской области (лесосстепь), 3 – Семиозерный лесхоз Кустанайской области (сухая степь).
Объекты и методы исследования
Сухоложский лесхоз расположен в 120 км к востоку от Екатеринбурга (570 с.ш., 620 в.д.). Территория лесхоза относится к сосново-березовому предлесостепному округу Зауральской равнинной провинции Западно-Сибирской равнинной лесорастительной области (Колесников и др., 1974). Cредняя температура +0,300С, абсолютный максимум +370С, абсолютный минимум -460С. Годовое количество осадков 510 мм. Длительность вегетационного периода 109 дней. Сосняки Iа-I классов бонитета составляют 44 %, а разнотравный тип леса - 55% покрытых лесом площадей. Лесные культуры занимают площадь около 10 тыс. га, из них 90 % площадей приходится на сосну. Лесистость территории 45 %. Средняя полнота сосняков 0,74, средний возраст 68 лет (Терин, 2004). Наши исследования выполнены на 5 пробных площадях в возрастном диапазоне сосны от 15 до 32 лет в разнотравном типе леса (Ia- II классы бонитета).
Саргатский лесхоз расположен в левобережье Иртыша в 110 км к северу от г. Омска (55030’с.ш., 73030’в.д.).
Территория лесхоза относится к Ишимо-Тюменскому сосново-березовому лесостепному округу, входящему в зону остепненных лесов (Крылов, 1959). Cредняя температура +0,280С, абсолютный максимум +380С, абсолютный минимум -400С. Годовое количество осадков 400 мм. Длительность вегетационного периода 160 дней. Лесистость 15,3%. Основные лесообразующие породы – береза и осина, занимающие соответственно 57 и 14 % площади лесных земель
Сосна представлена лишь культурами, которыми занято 29 % площади лесных земель, средний класс бонитета культур - II, полнота – 0,7. Рельеф равнинный, с незначительным превышением над уровнем моря (100-120 м). По гривам распространены серые лесные почвы, по западинам – солоди и солонцы, занимающие соответственно 60, 34 и 6 % лесных земель. Исследования выполнены на 9 пробных площадях, заложенных по двум градиентам - возрастному и эдафическому.
Семиозерный лесхоз расположен в Аман-Карагайском бору (52020’с.ш., 640 в.д.), который относится к провинции Тоболо-Убаганской равнинной степи степной зоны Тургайского прогиба и занимает площадь 80,9 тыс. га (Грибанов и др., 1970). Провинция расположена между Зауральским плато на западе, Тургайским плато на юге, левобережьем Ишима на востоке и колочной лесостепной равниной на севере. Основной породой является сосна обыкновенная, преобладающие типы почв - дерново-боровые и темно-каштановые.
Годовое количество осадков – 240 мм, влажность воздуха в летний период 25-30 %. Климат района резко континентальный. Среднемесячная температура июля составляет +200С, а января -180С. Годовая амплитуда абсолютных температур воздуха равна 890, среднегодовая температура +1,60. Вегетационный период длится 199 дней (Агроклиматический справочник…, 1969). В целом лесорастительные условия для сосны Аман-Карагайского бора очень жесткие. Недостаточное количество выпадающих атмосферных осадков в сочетании с высокими температурами воздуха в течение всего вегетационного периода и периодически повторяющимися засухами, довольно глубокое залегание грунтовых вод и небольшая зона капиллярного поднятия создают неблагоприятные условия для произрастания древесной растительности. Исследования выполнены на 8 пробных площадях, заложенных по двум градиентам - возрастному и эдафическому.
Подбор участков и работа на пробных площадях в трех таксонах проводились согласно нашим методическим разработкам (Усольцев, Залесов, 2005).
Результаты исследования и обсуждение
Характеристика пробных площадей, заложенных по возрастным и эдафическим градиентам, приведена в таблице 1. Для установления степени достоверности различия биопродуктивности древостоев культур сосны в трех таксонах принят метод регрессионного анализа зависимости фитомассы и первичной продукции от массоопределяющих показателей.
На первом этапе в трех таксонах выполнен сравнительный анализ биопродуктивности культур сосны высшей производительности (Iа-I класс бонитета). Вследствие невозможности в реальных условиях подобрать возрастные градиенты пробных площадей таким образом, чтобы возрасты во всех таксонах совпадали, мы сопоставляем не отдельные насаждения, а их возрастные тренды, т.е. зависимости показателей биопродуктивности от возраста, согласно модели:
lnPa и lnZa = a0 +a1 lnA + a2 (lnA)2, (1)
где Pa и Za - соответственно надземная фитомасса и годичная первичная продукция, т/га; А – возраст древостоя, лет.
Однако сопоставление культур высшей производительности трех таксонов по показателям биопродуктивности по уравнениям их зависимости лишь от возраста некорректно, поскольку густоты закладки культур в исследуемых таксонах разные. Соответственно различаются и текущие густоты культур (см. таблицу 1).
Достоверность и величину различия густоты культур в разных таксонах мы установили путем расчета зависимостей густоты от возраста, сопоставленных по таксонам посредством включения в модель блоковых фиктивных переменных (Дрейпер, Смит, 1973):
lnN = 6,4016 -2,4733 lnA + 0,3259 (lnA)2 -0,5641 X1 -1,1758 X2, (2)
где N– текущая густота культур, тыс. экз/га; X1 и X2 – блоковые фиктивные переменные, кодирующие принадлежность культур к тому или иному таксону следующим образом: сухая степь - X1 =0, X2 =0; предлесостепь - X1=1, X2=0; лесостепь - X1 =0, X2 =1. Коэффициент детерминации R2 уравнения (2) равен 0,783. Значимость констант при X1 и X2 в уравнении (2) составила по Стьюденту на уровне t05 соответственно 2,2 и 4,2, что выше нормативного значения t05 =2,0. Путем табулирования (2) по значениям возраста 10, 20, 30 и 40 лет установлено, что в Сухоложском и Саргатском лесхозах (соответственно в предлесостепи и лесостепи) текущая густота культур соответственно в 1,8 и 3,2 раз ниже, чем в Семиозерном (в сухой степи).
Поэтому в сравнительную модель кроме возраста насаждения ввели вторую независимую переменную - его текущую густоту, а также – блоковые фиктивные переменные, и регрессионная модель (1) приняла вид:
lnPа и lnZа = a0 +a1 lnA + a2 (lnA)2 +а3 (lnN)2 + a4Х1 + a5 X2. (3)
Коэффициенты детерминации составили для надземной фитомассы и надземной годичной продукции соответственно 0,927 и 0,436, т.е. уравнения (3) объясняли от 93 до 44 % изменчивости фитомассы и ее годичной продукции. Константы при блоковых фиктивных переменных Х1 и X2 оказались статистически не значимыми, поскольку соответствующий критерий Стьюдента варьировал от 0,45 до 1,50, что ниже нормативного, равной 2,0. Поэтому они исключены из (3), и рассчитаны уравнения, общие для трех таксонов
lnPа = -6,6698 +5,1028 lnА -0,5446 (lnA)2 + 0,2207 lnN, (4)
lnZа = 1,8127 +0,6290 lnA -0,1313 (lnA)2 -0,3592 lnN. (5)
Таким образом, различие трех таксонов по биологической продуктивности культур сосны, произрастающих по зональному градиенту в диапазоне от предлесостепи до сухой степи в возрасте от 10 до 50 лет, статистически не достоверно, как с учетом только возраста насаждений, так и с учетом двух факторов - возраста и густоты.
На втором этапе сопоставлены эдафические ряды биопродуктивности 20-летних культур (см. табл. 1), произрастающих в лесостепи (Саргатский лесхоз) и сухой степи (Семиозерный лесхоз), путем расчета регрессионной модели
lnPа и lnZа = a0 +a1 lnН20 + а2 lnN + a3Х, (6)
где Н20 – средняя высота древостоя в возрасте 20 лет; Х=0 для сухой степи и Х=1 для лесостепи. Однако значимость констант a3 при Х оказалась статистически не достоверной: t= 1,7 <t05 =2,0 для Pа ; t= 0,9 <t05 =2,0 для Zа. Поэтому рассчитаны обобщенные для экотона регрессии:
lnPа = 0,4142 +1,6472 lnН20 + 0,1594 lnN, (7)
lnZа = -0,8810 +1,6668 lnН20 -0,5745 lnN. (8)
Чтобы установить степень перекрытия показателей биопродуктивности культур на экотоне в петле гистерезиса необходимо привести (7) и (8) к густотам, характерным для того и другого таксона. Как уже было показано выше, показатели густоты по возрастным трендам для лесостепи и сухой степи существенно различаются. Для выявления степени достоверности различий по эдафическим градиентам рассчитано уравнение
N= 4,65 – 2,54 Х; R2 0,677, (9)
в котором константа при Х значима на уровне t05 , т.е. t = 3,5 > 2,0.
Таким образом, согласно (9) средние для эдафических градиентов густоты составили: для сухой степи 4,65 и для лесостепи 2,11 тыс. экз/га, т.е. в Саргатском лесхозе (лесостепь) текущая густота культур в 2,2 раза ниже, чем в Семиозерном (сухая степь).
После подстановки расчетных значений N (4,65 и 2,11)в (7) и (8) получили для лесостепи
lnPа = 0,5332 +1,6472 lnН20, (10)
lnZа = -1,3100 +1,6668 lnН20 (11)
и для сухой степи
lnPа = 0,6592 +1,6472 lnН20, (12)
lnZа = -1,7639 +1,6668 lnН20. (13)
Уравнения (10)-(13) протабулированы по значениям Н20, в соответствии с классами бонитета: для Ia, I, II, III и IV классов согласно шкале М.М. Орлова средние высоты Н20 равны 11,0; 8,5; 6,5; 5,5 и 3,5 м. Результаты табулирования, представленные графически (рисунок 3), дают представление о величине зон перекрытия петли гистерезиса в 20-летних культурах сосны на экотоне лесостепь-сухая степь по показателям класса бонитета, надземной фитомассы и годичной первичной продукции.
Заключение
Таким образом, несмотря на снижение уровня осадков в зональном направлении с 510 до 240 мм и повышение средней годовой температуры +0,3 до +1,60С, биологическая продуктивность культур в лучших условиях произрастания трех таксонов достоверно не различается согласно принципу взаимозаменяемости экологических факторов: понижение трофности почвы компенсируется повышением эдафического увлажнения и наоборот.
Таблица 1 - Таксационные показатели и биологическая продуктивность культур сосны на пробных площадях, заложенных на бореальном экотоне
№ пробной
площади |
Возраст, лет |
Средние |
Густота,
тыс. экз.
на 1га |
Запас, м3/га |
Класс
бонитета |
Фитомасса, т/га |
Годичная первичная продукция, т/га |
диаметр,
см |
высота, м |
Ствол
в коре |
Кора
ствола |
Ветви |
Хвоя |
Итого |
Ствол
в коре |
Кора
ствола |
Ветви |
Хвоя |
Итого |
Сухоложский лесхоз, предлесостепь, разнотравный тип леса (возрастной ряд) |
1 |
15 |
7,2 |
5,55 |
2,592 |
34,8 |
I |
11,5 |
2,16 |
6,49 |
5,98 |
24,0 |
1,36 |
0,217 |
1,27 |
2,33 |
4,96 |
2 |
18 |
7,4 |
7,71 |
6,045 |
107 |
I |
36,8 |
6,05 |
8,09 |
10,6 |
55,5 |
3,06 |
0,438 |
1,50 |
3,28 |
7,84 |
3 |
26 |
10,0 |
8,60 |
3,396 |
166 |
II |
42,8 |
5,72 |
12,8 |
10,4 |
66,0 |
2,59 |
0,310 |
1,67 |
3,48 |
7,75 |
4 |
29 |
12,1 |
12,2 |
2,733 |
207 |
I |
71,1 |
7,21 |
10,7 |
8,48 |
90,3 |
3,22 |
0,215 |
1,57 |
3,36 |
8,15 |
5 |
32 |
12,5 |
16,6 |
3,944 |
452 |
Iа |
147,2 |
13,5 |
18,1 |
9,39 |
174,7 |
3,66 |
0,299 |
1,92 |
2,92 |
8,50 |
Саргатский лесхоз, лесостепь, черноземы выщелоченные и осолоделые (возрастной ряд) |
1 |
10 |
5,7 |
4,5 |
2,264 |
17,9 |
Ia |
6,14 |
2,03 |
2,06 |
3,38 |
11,6 |
1,19 |
0,194 |
0,723 |
1,69 |
3,60 |
2 |
21 |
11,0 |
10,4 |
2,945 |
143 |
Ia |
48,9 |
5,84 |
10,4 |
7,16 |
66,4 |
5,48 |
0,521 |
1,448 |
2,88 |
9,81 |
3 |
30 |
15,6 |
14,8 |
1,925 |
282 |
Ia |
88,3 |
8,71 |
8,23 |
8,04 |
104,6 |
6,99 |
0,552 |
1,676 |
2,69 |
11,36 |
4 |
40 |
15,6 |
16,6 |
1,836 |
318 |
I |
110,2 |
7,81 |
11,9 |
10,6 |
132,7 |
4,54 |
0,322 |
0,817 |
5,33 |
10,69 |
5 |
50 |
18,4 |
18,7 |
1,603 |
289 |
I |
113,4 |
7,81 |
12,5 |
9,33 |
135,3 |
3,79 |
0,265 |
0,694 |
3,34 |
7,82 |
Саргатский лесхоз, лесостепь, черноземы выщелоченные и осолоделые (эдафический ряд) |
6 |
20 |
10,1 |
10,2 |
2,636 |
132 |
Ia |
40,86 |
4,30 |
12,66 |
10,58 |
64,10 |
5,49 |
0,578 |
1,283 |
4,24 |
11,01 |
7 |
20 |
12,3 |
8,9 |
1,873 |
88,9 |
I |
37,78 |
4,30 |
18,78 |
16,03 |
72,59 |
4,61 |
0,525 |
1,768 |
6,24 |
12,62 |
8 |
20 |
9,9 |
6,8 |
2,129 |
50,9 |
II |
22,29 |
2,84 |
9,05 |
8,48 |
39,82 |
2,75 |
0,350 |
0,776 |
3,08 |
6,61 |
9 |
20 |
8,2 |
5,1 |
1,794 |
27,1 |
III |
10,76 |
1,49 |
2,90 |
8,85 |
22,51 |
1,10 |
0,152 |
0,295 |
2,85 |
4,25 |
Семиозерный лесхоз, сухая степь, влажный бор, темно-каштановые почвы (возрастной ряд) |
1 |
10 |
3,3 |
4,0 |
18,87 |
52,3 |
I |
17,5 |
3,63 |
3,56 |
5,97 |
27,0 |
3,57 |
0,44 |
0,98 |
0,38 |
4,93 |
2 |
19 |
9,0 |
9,5 |
7,000 |
234 |
Ia |
81,3 |
11,4 |
13,8 |
12,8 |
107,9 |
2,23 |
0,11 |
2,78 |
3,71 |
8,72 |
3 |
23 |
11,0 |
10,2 |
3,830 |
203 |
I |
65,1 |
9,12 |
10,8 |
13,4 |
89,3 |
1,55 |
0,08 |
1,51 |
4,65 |
7,71 |
4 |
26 |
8,8 |
9,9 |
6,080 |
195 |
I |
69,3 |
10,2 |
6,36 |
5,77 |
81,4 |
0,19 |
0,06 |
0,93 |
1,71 |
2,83 |
Семиозерный лесхоз, сухая степь, влажный бор, , темно-каштановые* и дерново-боровые** почвы (эдафичекий ряд) |
15 |
20 |
10,8 |
9,3 |
3,850 |
191 |
I* |
63,6 |
7,40 |
12,7 |
10,9 |
87,2 |
1,57 |
0,10 |
1,90 |
3,83 |
7,30 |
16 |
20 |
7,0 |
7,0 |
6,330 |
100 |
II* |
34,0 |
5,75 |
5,80 |
6,11 |
45,9 |
0,95 |
0,07 |
0,94 |
1,84 |
3,73 |
17 |
20 |
7,7 |
5,5 |
3,240 |
55,2 |
III** |
19,0 |
3,88 |
6,41 |
5,35 |
30,8 |
0,60 |
0,07 |
0,80 |
1,94 |
3,34 |
18 |
20 |
6,1 |
4,8 |
5,180 |
48,1 |
IV** |
18,6 |
3,61 |
4,62 |
4,33 |
27,5 |
0,45 |
0,05 |
0,55 |
1,40 |
2,40 |
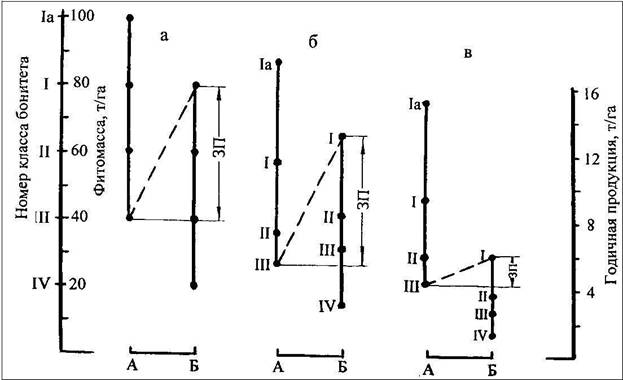
Рисунок 3 - Зоны перекрытия (ЗП) петли гистерезиса в 20-летних культурах сосны на экотоне лесостепь-сухая степь по показателям: (а) класса бонитета, (б) – надземной фитомассы, т/га, (в) – годичной первичной продукции, т/га; А – Саргатский лесхоз, лесостепь, Б – Семиозерный лесхоз, сухая степь
В то же время средние показатели продуктивности в том же направлении (с севера на юг) снижаются. Это следует из сопоставления соответствующих показателей, усредненных по эдафическим градиентам лесостепи и сухой степи (см. рисунок 3): класс бонитета снижается с I,5 до II,5; надземная фитомасса – с 53,0 до 38,7 т/га и годичная продукция – с 8,7 до 3,6 т/га.
Подтвержден выявленный в сосняках Окско-Мокшинского полесья (Коломыц и др., 1993) феномен раздвоения ландшафтно-экологического оптимума и наличия соответствующей зоны перекрытия биопродуктивности. Если в полесье раздвоение имело место в экотоне южная тайга-широколиственные леса, то в нашем исследовании - на рубеже лесостепи и сухой степи.
Зона перекрытия петли гистерезиса в 20-летних культурах сосны на экотоне лесостепь-сухая степь, обусловленная варьированием локальных почвенно-гидрологических условий,составила: по добротности местопроизрастания – два класса бонитета, с I по III (см. рисунок 3а), по надземной фитомассе – 37,4 (от 65,6 до 28,2) т/га (см. рис.3б) и по годичной первичной продукции – 1,5 (от 6,1 до 4,6) т/га (см. рисунок 3в). Относительная величина зоны перекрытия по годичной продукции (10%) значительно ниже, чем по фитомассе (42%), что, возможно, объясняется тем, что уже начиная с возраста 20 лет имеет место снижение энергии роста культур в условиях сухой степи.
Библиографический список
1. Агроклиматический справочник Кустанайской области (Под ред. Э.С. Зарембо). Алма-Ата, 1969. 200 с.
2. Алехин В.В. Объяснительная записка к геоботаническим картам (современной и восстановленной) бывшей Нижегородской губернии. Л., 1935.
3. Арманд А.Д. Гомеостазис экосистем // Экосистемы в критических состояниях. М.: Наука, 1989. С. 10-23.
4. Арманд А.Д., Ведюшкин М.А. Триггерные геосистемы (препринт). М.: Ин-т географии АН СССР, 1989. 51 с.
5. Арманд А.Д., Кушнарева Г.В. Переход экосистем через критические состояния в пространстве //Экосистемы в критических состояниях. М.: Наука, 1989. С. 75-138.
6. Базилевич Н.И., Гребенщиков О.С., Тишков А.А. Географические закономерности структуры и функционирования экосистем. М.: Наука, 1986. 297с.
7. Богатов А.О степной растительности в связи с вопросом о причинах безлесия южно-русских степей // Лесной журн. 1899. № 4. С. 550-575.
8. Будыко М.И. Изменение климата. Л.: Гидрометеоиздат, 1974. 280 с.
9. Варлыгин Д.Л., Базилевич Н.И. Связи продукции зональных растительных формаций мира с некоторыми параметрами климата // Изв. АН СССР. Сер. географ. 1992. № 1. С. 23-32.
10. Ведюшкин М.А. Моделирование пространственных переходов между фитоценозами // Математическое моделирование популяций растений и фитоценозов. М.: Наука, 1992. С. 24-30.
11. Габеев В.Н.Экология и продуктивность сосновых лесов. Новосибирск: Наука, 1990. 229 с.
12. Горбатенко В.М. Влияние основных климатических факторов на первичную биологическую продуктивность сосновых фитоценозов // Стационарные гидрологические исследования в лесах Сибири. Красноярск: ИЛиД СО АН СССР, 1975. С. 212-220.
13. Грибанов Н.И. Подгородная лесная дача Сибирской сельскохозяйственной академии // Тр. Сибирской с.-х. академии. Т. 11. Омск, 1923. С. 3-14.
14. Грибанов Л.Н., Лагов И.А., Чабан П.С. Леса Казахстана Т. 5. // Леса СССР. М.: Наука, 1970. С. 5-76.
15. Докучаев В.В. Методы исследования вопроса: были ли леса в южной степной России? // Тр. Вольного эконом. общ-ва. 1889. № 1. С. 1-38.
16. Дрейпер Н., Смит Г. Прикладной регрессионный анализ.- М.: Статистика, 1973. 392 с.
17. Завалишин А.А. Почвы Кузнецкой лесостепи // Труды СОПС Акад. Наук. Серия сибирская. Т. 20. 1936. С. 165.
18. Колесников Б.П., Зубарева Р.С., Смолоногов Е.П. Лесорастительные условия и типы лесов Свердловской области. Свердловск: УНЦ АН СССР, 1974. 176 с.
19. Коломыц Э.Г. Опыт теоретико-множественного моделирования высокогорных экосистем // Тр. ВГИ. 1984. Вып. 58. Л.: Гидрометеоиздат. С. 65–69.
20. Коломыц Э.Г., Юнина В.П., Сидоренко М.В., Воротников В.П. Экосистемы хвойного леса на зональной границе: Организация, устойчивость, антропогенная динамика. Нижний Новгород: Ин-т экологии Волжского бассейна РАН, 1993. 347 с.
21. Коржинский С.И. Северная граница черноземной области восточной полосы Европейской России в ботанико-географическом и почвенном отношениях // Тр. Общества естествоиспытателей Казанского ун-та. 1888. Т. 18. № 5. 253 с. (Часть 1); 1891. Т. 22. № 6. 201 с. (Часть 2).
22. Костычев П.А. Почвы черноземной области России, их происхождение, состав и свойства. С.-Пб., 1886. 124 с.
23. Кофман Г.Б., Кузьмичев В.В. Видоспецифичность естественного изреживания древостоев // Формирование эталонных насаждений. Ч. 1. Каунас, Гирионис: ЛитНИИЛХ, 1979. С. 99-101.
24. Краснов А.Н. Материалы для флоры Полтавской губернии // Тр. Общ-ва испытателей природы при Харьковском ун-те. 1890. Т. 24. С. 446.
25. Крылов Г.В. Лесорастительное районирование Сибири // Изв. Томск. отд-ния Всерос. ботан. общ-ва. Т. 4. Новосибирск, 1959. С. 115-149.
26. Лавренко Е.М. Степи СССР // Растительность СССР. Т. 2. М.; Л.: Изд-во АН СССР, 1940. С. 1-265.
27. Леса России – 2005 // Российская лесная газета. 2006. № 8-10.
28. Майков А. Заметки на географию древней Руси // Журн. Министерства народного просвещения. 1874. № 8. С. 259-276.
29. Митрофанов С.В. Состояние культур кедра сибирского в лесостепи Южного Урала // Леса Урала и хоз-во в них. Вып. 27. Екатеринбург: УГЛТУ, 2006. С. 179-184.
30. Одум Юдж.. Основы экологии. Пер. с 3-го англ. изд. М.: Мир, 1975. 740 с.
31. Палимпсестов И. Степи юга России были ли искони веков степями и возможно ли облесить их? // Лесной журн. 1882. № 2. С. 93-141.
32. Пермский А. Нынешние культуры в Велико-Анадольском степном лесничестве // Лесной журнал. 1876. № 1. С. 1-14. № 2. С. 1-21.
33. Раменский Л. Г. Введение в комплексное почвенно-геоботаническое исследование земель. М.: Сельхозгиз, 1938. 620 с.
34. Рупрехт Ф. Геоботанические исследования о черноземе. С.-Пб., 1866. 36 с.
35. Савич М. Опыт разведения леса в степи Уральского казачьего войска // Лесной журн. 1885. № 1. С. 22-35.
36. Собичевский В. Старинный вопрос, были ли в исторические времена леса в южной степной России, и новейшие его решения // Лесной журн. 1890. № 3. С. 333-350.
37. Сукачев В.Н. Избр. тр. В 3 т. Т. 1. Основы лесной типологии и биогеоценологии. Л.: Наука, 1972. 418 с.
38. Танфильев Г.И. Пределы лесов на юге России. С.-Пб., 1894. 174 с.
39. Таран И.В. Сосновые леса Западной Сибири. Новосибирск: Наука, 1973. 292 с.
40. Терин А.А.Состояние сосновых насаждений и перспективы их хозяйственного использования после подсочки в Среднем Зауралье: Автореф. дис… канд. с.-х. наук. Екатеринбург: УГЛТУ, 2004. 21 с.
41. Усольцев В.А. Фитомасса лесов Северной Евразии: база данных и география. Екатеринбург: УрО РАН, 2001. 707 с.
42. Усольцев В.А., Залесов С.В. Методы определения биологической продуктивности насаждений. Екатеринбург: УГЛТУ, 2005. 147 с.
43. Уткин А.И. О возможной динамике лесной растительности в экотонах Северной Евразии при глобальном потеплении // Классификация и динамика лесов Дальнего Востока: материалы международной конференции. Владивосток, 2001. С. 125 – 127.
44. Фрейдин. Богата ли Сибирь ценными лесами? // Лесной журн. 1900. № 6. С. 799-813.
45. Чернов Н.Н. Лесные культуры на Урале. Ч.1. Екатеринбург: УГЛТА, 1998. 541 с.
46. Чиллингуорт Д. Структурная устойчивость математических моделей. Значение методов теории катастроф // Математическое моделирование. Под ред. Дж. Эндрюса и Р. Мак-Лоуна. М.: Мир, 1979. С. 249-276.
47. Шиятов С.Г., Терентьев М.М., Фомин В.В. Пространственно-временная динамика лесотундровых сообществ на Полярном Урале // Экология. 2005. № 2. С. 1-8.
48. Bar K. Die uralte Waldlosigkeit der sudrussischen Steppe // Beitrage zur Kenntniss des russischen Reiches. 1856. Bd. 18. S. 109-115.
49. Bugmann H., Pfister C. Impacts of interannual climate variability on past and future forest composition // Reg. Еnviron. Change. 2000. № 1. Р. 112–125.
50. Korner Ch. A re-assessment of high elevation treeline positions and their explanation // Oecologia. 1998. Vol. 115. P. 445-459.
51. Middendorf A.F. Sibirische Reise. 1864. Bd. IV. Teil I. S. 613-728.
|


