|
Активизация патогенных свойств грибов комплекса Armillaria mellea sensu lato в хвойных лесах юга Восточной Сибири
Павлов И.Н., Миронов А.Г., Юшкова Т.Ю.
В последнее десятилетие на территории южной тайги в границах Красноярского края отмечена интенсивная куртинная гибель сосны обыкновенной, ели сибирской, пихты сибирской. Усыханию подвержены деревья всех классов Крафта. Погибают экземпляры, не испытывающие каких-либо признаков угнетения, с хорошо развитой кроной. По комплексу диагностических признаков к основной причине гибели отнесены виды, входящие в комплекс Armillaria mellea s.l. (A. borealis, A. ostoyae). Наибольшая вредоносность опенка установлена в спелых и приспевающих сосняках сформировавшихся на маломощных почвах, подстилаемых твердыми горными породами, после рубок главного пользования и следующих за этим пожаров, находящихся в зоне хронического атмосферного загрязнения и интенсивного рекреационного влияния. Смена коренных типов леса и антропогенное воздействие являются первичными факторами снижения устойчивости. Глобальное изменение климата, создавая более благоприятные условия для развития болезней и вредителей, способствует росту поражающего биотического воздействия на растения. Указанные факторы являются определяющими для активации механизма разрушения древостоя включающего следующие этапы: дальнейшее ослабление отдельных деревьев в результате напряженной внутриценотической конкуренции, воздействия рака-серянки > рост вирулентности и агрессивности опенка > значительно ослабленные корневыми патогенами деревья не могут сопротивляться стволовым вредителям (малому и большому сосновым лубоедам) в период их массового размножения и погибают. При отсутствии воздействия лубоедов последствия армилляриоза не были бы столь значительны и скоротечны.
Over the last decade intensive clump death of Pinus sylvestris, Picea obovata, Abies sibirica has been observed in the territory of south taiga within the boundaries of Krasnoyarsk Region. Trees of all Kraft classes are subjected to drying. Specimens with well-developed crown, showing no signs of suppression, perish. By the complex of diagnostic characters, species belonging to the complex Armillaria mellea s.l. (A. borealis, A. ostoyae) have been attributed to the principal cause of death. The highest injuriousness of Armillaria has been ascertained in mature and ripening pine forests formed at shallow soils underlied by hard rock, after major harvests and subsequent fires, located in the zone of chronic atmospheric pollution and intense recreational impact. Change in aboriginal forest types and man impact are the primary factors of decrease in stability. Global climate change, creating more favourable conditions of development of diseases and pests, contributes to increase in damaging biotic effect on plants. The specified factors are determinative for destruction mechanism of the stand; the mechanism includes the following stages: further weakening of separate trees due to tense intra-coenotic competition, impact from resin canker> increase in Armillaria virulence and aggressivity > trees considerably weakened by Armillaria cannot withstand trunk pests (Tomicus minor and Tomicus piniperda) in the period of their mass propagation, and die.
Колебания климата, интенсификация антропогенного воздействия, межрегиональ-ное перемещение болезней и вредителей являются причиной ухудшения санитарного состояния лесов (Алексеев и др., 1999; Слободян и др., 2001; Стороженко, 2001; Матусевич, 2003; Vertui, Tagliaferro, 1998; Bruhn et al., 2000; Hogg et al., 2002). Отмечаются случаи быстрого усыхания как отдельных вполне здоровых деревьев и разновозрастных куртин, так и древостоев в целом. При этом часто первопричина не может быть установлена. Имеют место появление новых по симптомам поражения (ранее не отмечаемых в регионе) случаев заболевания деревьев. Если ранее некоторые болезни представляли особую опасность для искусственно созданных насаждений или ослабленных, то в последние десятилетия отмечаются случаи их патогенного воздействия на казалось бы абсолютно здоровые насаждения. К числу таких заболеваний может быть отнесен и армиллариоз (заболевание вызываемое грибами комплекса Armillaria mellea s.l.).
В комплекс входит несколько видов, отличающихся по морфологии, экологии и патогенности. К. Корхонен (1978) в результате скрещивания гаплоидных чистых культур (mating tests), основанного на методе сексуальной несовместимости разных видов, в структуре комплекса A. mellea s.l. на территории Западной Европы, учитывая морфологические и анатомические различия базидиом, выделил несколько интрафертильных групп A (A. borealis Marxm. & Korhonen), B (A. cepistipes Velen.), C (A. ostoyae (Romagn.) Herink), D (A. mellea (Vahl: Fr.) Kumm.), E (A. gallica Marxm. & Romagn.).
Исторически армилляриоз рассматривал-ся как заболевание ослабленных деревьев (Shaw III, Kile, 1991). Только в последние годы некоторые виды опенка получили признание как первичные патогены в естественных лесах (Shaw III, Kile 1991; Baumgartner, Rizzo, 2001; Schwarze, Ferner, 2003).
В то же время в России и Белоруссии остается преобладающим мнение об опенке как о вторичном патогене, способном заражать лесные культуры и деревья, произрастающие в стрессовых условиях (Соколов 1964; Федоров, 1984; Иванов, 1981; Селочник, 1992; Звягинцев, 2003)
Н.Н. Селочник и Н.К. Кондрашева (1991) отмечают, что, несмотря на большой потенциал инфекции опенка в пойменных и нагорных дубравах, внедрение патогена под кору ствола и корней происходит в основном у усыхающих и свежеусохших деревьев, реже – у сильно ослабленных.
В работе C. Shaw III, G. Kile (1991) показана высокая патогенность А. mellea и А. ostoyae (другие виды данного рода – слабые патогены) и зависимость поражения от благоприятного для опенка гидро-термического режима (физиологический стресс хозяина нарушает баланс устойчивости в пользу патогена).
Отмечается различная вредоносность одного и того вида опенка в различных регионах. Так, в США A. mellea проявляет сильную патогенность, вызывая деградацию дубовых лесов и гибель хвойных в Калифорнии, в горных же лесах Миссури этот вид оказался вирулентным только по отношению к ослабленным соснам (Bruhn et al., 2000, Baumgartner, Rizzo, 2001). С возрастом деревья оказываются более устойчивыми, снижая способность патогена вызывать заболевание (Robinson, Morrison 2001).
В целом наибольший вред A. ostoyae, как первичный патоген причиняет в освоенных лесах умеренного климата центральной Европы и западной части северной Америки (наиболее чувствительны культуры хвойных до 15-30 лет) (Wahlsrom, 1992; Korhonen, 2004). Деревья сосны старших возрастов оказываются более устойчивыми к заболеванию.
Сведения о высокой патогенности опенка в таежной зоне России отсутствуют. В хвойных насаждениях Урала и Сибири опенок - типичный представитель спелых и перестойных насаждений и вызывает незначительный отпад ели, пихты, кедра и лиственницы (Жуков, 1978). При лесопатологическом обследовании высокогорных темнохвойных лесов грибы комплекса A. mellea занимают одно из последних мест, рассматриваясь главным образом как сапротрофы. Их присутствие ассоциируется с сухостоем и пнями березы и ольхи кустарниковой и очень редко с основаниями стволов усыхающих пихт и елей (Петров, 1989).
Программа, методика и объекты
В нижнем течении р. Караульной (южная тайга) на прилегающих сопках в последнее десятилетие наблюдается интенсивная куртинная гибель сосны обыкновенной, ели сибирской, пихты сибирской. Усыханию подвержены деревья всех классов Крафта. Погибают экземпляры, не испытывающие каких-либо признаков угнетения, с хорошо развитой кроной. Продолжительность жизни хвои, ее размеры, прирост побегов не отличаются от средних значений по древостою. При этом гибель наступает очень быстро. В ряде случаев отмечена гибель после образования текущего прироста с несформировавшейся хвоей. По комплексу диагностических признаков к основной причине гибели отнесены виды, входящие в комплекс A. melleas.l.
Проведенный нами анализ многочисленных ключей (Павлов, Миронов, Кутафьева, 2006) к определению видов Armillaria, разработанных для европейских стран, позволил выявить безусловные отличительные морфологические признаки базидиом. При их использовании были установлены виды, произрастающие на исследуемой территории: A. borealis, A. cepistipes, A. ostoyae.
Процессы интенсивного изреживания носят ярко выраженные пространственные закономерности. Каждый очаг обычно начинается с инфицирования одного, реже нескольких деревьев, от которых опенок распространяется более или менее равномерно во все стороны, вызывая поражение здоровых деревьев и последующее их отмирание (Негруцкий, 1986; Соколов, 1964). Следовательно, и форма пробной площади должна соответствовать биологии изучаемого процесса (имеющей кольцевой характер), т.е. быть овальной или круглой. Закладка круглой площади значительно проще, в свою очередь овал может быть сформирован путем последовательного заложения двух или более круговых площадок.
Применение круговых площадок нашло широкое применение при исследовании возобновления, для изучения фитогенного поля древесных пород (травостой описывается на квадратных площадках в 0.1 м2, располагающихся по трансектам, пересекающим подкроновое пространство по радиусу или по секущей (Самойлов, 1983)), при таксации лесосек (круговые реласкопические площадки, перечет на круговых площадках постоянного радиуса) (Наставление…, 1993). При количественной оценке структуры древостоя используется сочетание круговых учетных площадок (радиусом, равным максимальной длине главных латеральных корней деревьев) с размещением по центру квадратных площадок (Санникова, 1992). На квадратных площадках осуществляется учет подроста и факторов среды, определяется расстояние до деревьев, входящих в круговую площадку
Классические методы картирования предполагают разбивку исследуемой территории регулярной сеткой с шагом 2, 5 или 10 метров в зависимости от густоты древостоя (Емшанов, 1999). Небольшие размеры ячеек обеспечивают точное определение координат отдельных элементов древостоя. Исследования требуют больших трудозатрат, однако окупаются высокой степенью точности съемки.
Нами для снижения трудоемкости при сохранении точности предлагается использовать при таксации и картировании древостоя круговые пробные площади (ПП) переменного радиуса (где нет густого подроста и подлеска). За центр площади принимается место, обеспечивающее хорошую просматриваемость стволов окружающих деревьев и (или), при изучении кольцевых процессов – точка влияния (распространения). Способ хорошо зарекомендовал себя при изучении кольцевых процессов распространения агрессивных видов грибных возбудителей.
В центре инфекции, на месте погибших и взятых в качестве моделей деревьев устанавливается буссоль (для определения угла по лимбу между направлениями на С (0о) и на центр картируемого дерева) и шпилька с мерным шнуром (если не используется дальномер) (рисунок 1).
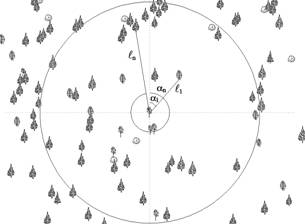
Рисунок 1- Фрагмент круговой площади:
? – угол, град; ? - расстояние от центра площади до дерева, м; 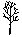 - сухостой сосны; - сухостой сосны; 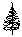 - сосна обыкновенная; - сосна обыкновенная;  - пень (в - пень (в 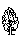 – береза повислая – береза повислая
Данные заносятся в таблицу. Представленная на рисунке схема построена автоматизировано по данным таблицы с помощью программного обеспечения, работающего в качестве расширения ESRI ArcMap. ГИС-приложение предназначено для хранения картографической информации с пробных площадей и автоматизации расчета таксационных показателей.
Геоинформационные технологии позволяют осуществлять любую сегментацию пробной площади. Для изучения процессов расширения очагов и последующего возобновления в «окнах» проведена оценка таксационных показателей по кольцам, различно удаленным от центра.
В случае если удаленное дерево не просматривается из-за рядом стоящего ствола, применяется способ определения горизонтальной поправки. В программном обеспечении предусмотрен пересчет координат с учетом угла, засеченного по видимой части мерного шеста и поправки.
Хорошей проверкой точности картирования является пересечение двух пробных площадей при формировании овальной ППП. Деревья, измеренные из разных центров и относящиеся одновременно к двум пробным площадям, при автоматизированной визуализации накладываются точно друг на друга.
В насаждениях с преобладанием сосны обыкновенной заложено три полигона площадью 10,8 га, 1,0 га и 0,18 га. Балл санитарного состояния, класс роста, диаметр на высоте 1,3 м измерялись у всех деревьев, высота – только у 35-50 экземпляров каждой породы, входящей в состав древостоя. На полигоне № 1 (10,8 га), заложено 13 круговых ППП общей площадью 1,17 га (без учета наложения – 1,36 га). В насаждениях с преобладанием пихты заложено 2 ППП; с преобладанием ели – 3.
В очагах усыхания у 80 погибших деревьев сосны на высоте 0,5-1,0 м были взяты поперечные спилы. В качестве контроля были выбраны особи сосны (30 шт.) без признаков нарушения жизнедеятельности, без следов болезней и вредителей. Год гибели деревьев установлен методом перекрестной датировки значений радиального прироста погибших деревьев с контрольными.
Наряду с описанием почвенных разрезов для изучения глубины корнеобитаемого слоя был изготовлен металлический щуп (по аналогии с «мечом Колесова»). Им в нескольких радиальных направлениях от центра на круговых пробных площадях было сделано более 600 измерений.
Для изучения влияния опенка на процессы естественного возобновления на различном удалении от центров куртинного усыхания (от 0 до 10 м, с шагом 2 м) произведена инвентаризация подроста. В качестве контроля были взяты примыкающие древостои без признаков заболевания, имеющие сходные таксационные и лесоводственные показатели.
обсуждение и анализ результатов
Не исключая наличия предрасполо-женности к заболеванию у ослабленных деревьев, следует учитывать и имеющие место глобальные изменения в окружающей среде (климат, антропогенное воздействие). Кроме всего прочего климатические особенности, а также позднее начало активной лесозаготовительной деятельности на территории Сибири, являются причиной некоторого отставания во времени проявления эпифитотий корневых патогенов от Европы и Северной Америки.
В таблице 1 показаны результаты анализа встречаемости видов Armillaria на изученной территории образования эпифитотия. Наиболее агрессивный патоген A. оstoyae доминирует на сосне обыкновенной (в т.ч. на живых деревьях с внедрением под кору).
Сухостой, образовавшийся за достаточно короткий период (5-8 лет) по круговым пробным площадям составляет от 5 до 33 %. По степени очагового поражения сосновые древостои на исследуемых полигонах (рисунок 2) имеют 3-4 балл по шестибальной шкале (по В.Г. Стороженко, 2001). Из хвойных деревьев наименее подвержена патогенному действию опенка пихта сибирская.
Распределение числа деревьев по диаметру является важным показателем, характеризующим состояние древостоя. Успешная аппроксимация распределения деревьев по диаметру в очагах корневых патогенов оказалась возможной только с использованием многокомпонентной смеси нормальных распределений (ряды распределения усохших деревьев имеют ярко выраженную положительную асимметрию, живых – отрицательную) (рисунок 3). Наличие сухостоя диаметром 16-46 см свидетельствует об интенсивном патогенном характере отпада. Первая составляющая в распределении живых деревьев по диаметру (Dср живых = 15,4 см) образована в результате отсутствия своевременного отпада отстающих в росте особей сосны (по генотипу или произрастающие на маломощных почвах). Среди них и образуется первая составляющая сухостоя со средним значением диаметра 11 см.
Отпад происходит естественно (действие патогенов вторично) в результате угнетения со стороны соседних деревьев. В дальнейшем именно на их основе происходит усиление вирулентности и агрессивности опенка. Поэтому технологические решения, как при естественном возобновлении, так и при искусственном восстановлении должны быть направлены на предотвращение образования значительно угнетенного яруса деревьев (редкое размещение посадочного материала).
Таблица 1 - Приуроченность видов Armillaria к древесным растениям-хозяевам
Вид растения-хозяина |
Балл санитарного состояния |
Встречаемость, % |
Вид Armillaria |
Сосна обыкновенная
Pinus sylvestris |
1 |
3 |
А. ostoyae |
2 |
10 |
3 |
4 |
5 |
14 |
6 |
3 |
Ель сибирская
Picea obovata |
1 |
1 |
А. cepistipes |
2 |
А. borealis |
2 |
4 |
5 |
6 |
6 |
2 |
А. ostoyae |
3 |
А. cepistipes |
Пихта сибирская
Abies sibirica |
2 |
3 |
А. borealis |
Береза повислая
Betula pendula |
2 |
7 |
5 |
А. cepistipes |
3 |
3 |
А. borealis |
6 |
4 |
1 |
А. ostoyae |
Осина
Populus tremula |
5 |
9 |
А. borealis |
1 |
А. ostoyae |
3 |
А. cepistipes |
6 |
8 |
4 |
А. borealis |
 
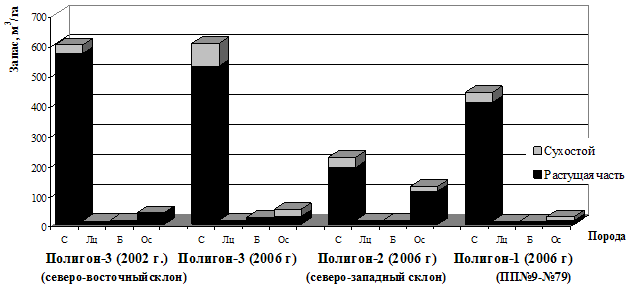
Рисунок 2 – Таксационная характеристика сосновых древостоев, имеющих очаговое поражение
Вторая составляющая отпада (Dср = 21 см) образована действием опенка, как первичного патогена, что способствует рассечению общего распределения живых деревьев по диаметру на две составляющие.

Рисунок 3 - Распределение числа деревьев сосны по диаметру в пораженных древостоях (1– растущая часть; 3 – сухостой) и в контроле (2 – растущая часть; 4 – сухостой)
В качестве контроля было выбрано сходное по таксационно- лесоводственным показателям насаждение без признаков заболевания, примыкающее к древостою с признаками армилляриоза. Естественный отпад в пределах нормы.
 В очагах развития армилляриоза установлена значительная гибель подроста (при наличии мицелия на корнях, редко – базидиом) (рисунок 4). Несмотря на то, что световой режим и отсутствие корневой конкуренции на месте выпавших деревьев - оптимальны для роста естественного возобновления, его смертность значительно превышает контроль. С удалением от центра – количество жизнеспособного подроста увеличивается. Это является надежным подтверждением патогенного действия опенка. В очагах развития армилляриоза установлена значительная гибель подроста (при наличии мицелия на корнях, редко – базидиом) (рисунок 4). Несмотря на то, что световой режим и отсутствие корневой конкуренции на месте выпавших деревьев - оптимальны для роста естественного возобновления, его смертность значительно превышает контроль. С удалением от центра – количество жизнеспособного подроста увеличивается. Это является надежным подтверждением патогенного действия опенка.
Анализ радиального прироста в очагах корневых патогенов выявил существенные различия в динамике (рисунок 5). Четко выделяется период 1920-1940 гг. В это время древостой сосны вступил после смыкания крон в фазу жесткой конкуренции и активной дифференциации. В связи с его одновозрастностью и достаточно равномерным размещением отдельных особей на площади результаты конкуренции неблагоприятно сказываются на всем древостое в целом. Происходит «эффект группового угнетения» (Погребняк, 1968; Романовский, 2002).

Рисунок 4 - Количество подроста сосны (в пересчете на крупный) на различном удалении от центра очага инфекции:
Это сопровождается снижением прироста у всех экземпляров взятых в качестве моделей (впоследствии все они погибли). Возможно, уже в тот период и произошло заражение армилляриозом. Первоначальное развитие инфекции произошло на пнях вырубленного древостоя. У деревьев взятых в качестве контроля в том же насаждении (без каких-либо признаков ослабления) отсутствует данный период депрессии. Это подтверждает высокую опасность длительного нахождения древостоя в условиях избыточной загущенности без активной дифференциации и своевременного отпада (часто имеет место на плодородных, но маломощных почвах при одновременном появлении возобновления).
По состоянию и закономерностям радиального прироста выделено пять групп деревьев (рисунок 5):
1. Деревья IV-V классов роста, испытывающие значительное ценотическое давление, с равномерным снижением прироста после смыкания крон (до 0,16 мм). Первичным фактором гибели является общее ослабление в результате конкуренции за свет. А. melleas.l. в данном случае выступает как вторичный паразит.
2. Устойчивое снижение прироста деревьев начинается с 1973 г. Все особи данной группы (II-III классы Крафта) имеют признаки поражения смоляным раком (в общем количестве погибших деревьев их было выделено до 26%). Смоляной рак выступает дополнительным фактором снижения устойчивости к корневым патогенам. В очагах деревья данной группы усыхают в первую очередь, наряду с представителями предыдущей группы. На данном этапе происходит рост агрессивности и вирулентности опенка.
3. Деревья, входящие в данную группу, имеют радиальный прирост ниже среднего по древостою и занимают промежуточное положение в пологе (II-III классы), не имеют признаков заболевания смоляным раком и погибают после образования очага и достаточного накопления инфекции.
4. Гибель с коротким (2-5 лет) предварительным снижением радиального прироста без образования поздней древесины. До периода угнетения роста различие с контрольными значениями не существенно. Стволы имеют высокую степень заселенности малым и большим сосновым лубоедами, ускорившими гибель ослабленных корневыми патогенами деревьев. Деревья 2 и 4 группы до 1978 г. развиваются синхронно. Далее, с развитием смоляного рака, радиальный прирост деревьев второй группы снижается более значительно.
5. Гибель без предварительного снижения радиального прироста на стадии образования поздней древесины. Дендрохронологический ряд практически не отличается от контрольных значений. Данные деревья (самые крупные в насаждении) произрастают свободно в окнах, в окружении старых пней (в основном лиственных пород) с образованием большого числа базидиом опенка, что способствует росту его патогенности. При хорошем прогревании почвы патоген активизируется в более ранние сроки и к концу вегетационного периода (при одновременном воздействии малого и большого сосновых лубоедов) способен привести инфицируемое им дерево к гибели (что объясняет факт образования поздней древесины в последний год жизни). Деревья, входящие в 4 и 5 группы, погибают после достаточного накопления инфекции в очаге.
В развитии любого патогена, в т.ч. и опенка необходимым условием развития вспышки заболевания является образование для него благоприятных условий при одновременном воздействии стрессовых факторов на хозяина (особенно на первом этапе развития болезни). Внешне большинство деревьев не имеют каких-либо симптомов угнетения. И если бы не очаговое усыхание, исследуемые древостои можно было бы отнести к здоровым. Для исследуемого района характерны маломощные почвы с высоко плодородным гумусовым горизонтом. Вероятно, для данных условий климаксовыми могут быть древостои с преобладанием пихты сибирской.

Рисунок 5 - Динамика радиального прироста сосны обыкновенной (к- контроль; 1-5 – группы деревьев, погибших в результате деятельности А. melleas.l; +,?,0, x, ^ – значения радиального прироста отдельных особей сосны в год гибели)
Практически все почвы исследуемых насаждений относятся к подтипу дерново-карбонатных типичных, для которых характерны малая мощность профиля, красно-бурая или коричневая окраска, многочисленные включения карбонатных пород (известкового щебня), вскипание от 10% раствора HCL в верхнем гумусовом горизонте, суглинистый гранулометрический состав с незначительным содержанием песчаной фракции (Почвы СССР, 1979; Горбачев, 1986). pH водной вытяжки всех исследуемых почвенных горизонтов колеблется в интервале от 6,4 до 7,2, т.е. близка к нейтральной реакции.
При анализе появления очагов усыхания сосны на различных элементах рельефа в сочетании с исследованием глубины корнеобитаемого слоя установлена устойчивая закономерность. На вершинах сопок, с очень мелким быстро пересыхающим корнеобитаемым слоем (крайне неблагоприятно для развития опенка), а также на глубоких почвах (высокая устойчивость хозяина) практически отсутствует гибель деревьев. Наиболее неблагоприятные для сосны условия складываются на неглубоких почвах (корнеобитаемый слой до 30 см) подстилаемых твердыми горными породами недоступными для корневой системы (рисунок 6). В данном случае сосна образует корневую систему с редуцированным стержневым корнем (установлено при многочисленной корчевке пней при строительстве дороги). В молодом возрасте насаждение развивается как высокопродуктивное. Далее, при превышении эдафического потенциала, наступают стрессовые условия снижающие устойчивость к А. melleas.l. Дополнительным необходимым условием развития болезни является умеренная влажность почвы. Очаги усыхания отсутствуют в местах, где осадки быстро уходят вниз по склону.
Размещение усыхающих деревьев на полигоне неравномерно. Сливаясь усыхающие куртины образуют очаги овальной формы до 1 га. (рисунок 7). Выделение осуществлено с помощью процедуры кластерного анализа. Характерная форма овала (вытянутость с северо-востока на юго-запад), возможно, определяется рельефом, господствующими ветрами, направлением водотока. Границы очагов не имеют четко выраженного характера.
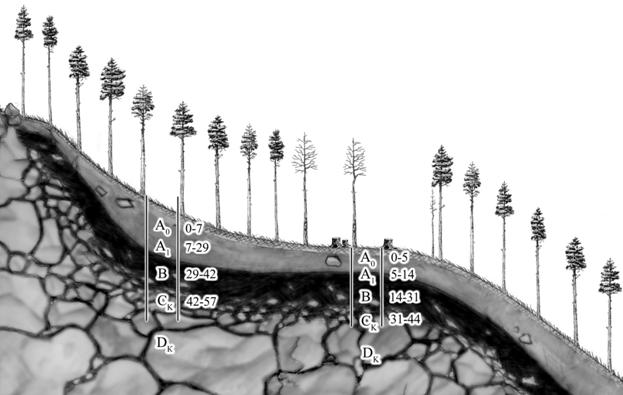
Рисунок 6 - Эдафические закономерности образования очагов усыхания
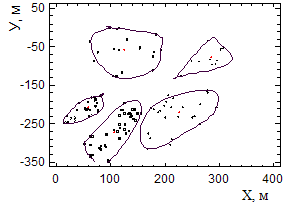
Рисунок 7 - Распределение погибших деревьев на полигоне (процедура кластерного анализа, Ward's Distance Metric: Squared Euclidean).
Безусловно, первичными факторами снижения устойчивости являются смена коренных типов леса и антропогенное воздействие. Так, исследуемые одновозрастные сосняки сформировались после рубок главного пользования и следующих за этим пожаров на маломощных почвах подстилаемых твердыми горными породами (как следствие образуется корневая система с редуцированным стержневым корнем) и находятся в зоне хронического атмосферного загрязнения и интенсивного рекреационного влияния.
Смена коренных типов леса и антропогенное воздействие являются первичными факторами снижения общей устойчивости лесных ценозов. Изменение климата, в свою очередь, создавая более благоприятные условия для развития болезней и вредителей, способствует росту поражающего биотического воздействия на растения. По данным Cредне-Сибирского межрегионального территориального управления по гидрометеорологии и мониторингу окружающей среды, для исследуемой территории характерен рост температуры приземного слоя воздуха, наиболее существенный в последние два десятилетия. Более теплая зима создает лучшие условия для зимовки вредителей и болезней. Значительный рост температуры в мае (сумма эффективных температур >5оС возросла на 90о С) обеспечивает более раннее начало роста опенка и увеличение продолжительности его вегетационного периода (рисунок 8).

Рисунок 8. – Изменение температуры приземного слоя воздуха на исследуемой территории
Указанные факторы являются определяющими для запуска многоэтапного механизма разрушения древостоя, где основная роль принадлежит опенку.
На первом этапе в сосновых древостоях с высокой равномерной полнотой (0,8-1,2) и без ярко выраженных куртин усохших деревьев отпаду подвергается сосна IV-V классов Крафта а также спелые и перестойные деревья осины и березы. Происходит накопление инфекции. Наличие плодовых тел опенка на некоторых экземплярах сухостоя, живых и усыхающих деревьев и макроскопические особенности гнили свидетельствуют о развитии очага инфекции.
На втором этапе происходит гибель деревьев образующих основной полог насаждения, ослабленных в результате напряженной внутриценотической конкурен-ции (эффект группового угнетения), воздействия рака серянки и других болезней и вредителей нелетального характера. В дальнейшем они становятся центром образования очагов усыхания.
На третьем этапе - лучшая прогреваемость почвы в разреженных насаждениях, интенсивное развитие живого напочвенного покрова, ослабление древостоя рекреационным и техногенным воздействием способствуют усилению вирулентности и агрессивности опенка. Условия, благоприятствующие заражению и массовому воспроизводству патогена, увеличивают вероятность образования более вирулентных рас (Помазков, 1990).
В образовавшихся окнах более вирулентный (чем первоначально) патоген, распространяясь от погибших деревьев через сросшиеся корни по многим направлениям, осуществляет воздействие на деревья I-II классов Крафта. Значительно ослабленные деревья не могут сопротивляться стволовым вредителям (малому и большому сосновым лубоедам) в период их массового размножения и погибают.
Библиографический список
- Алексеев, В.А. Состояние пихтовых лесов Кузнецкого Алатау / В.А. Алексеев, В.В. Астапенко, Ю.Г. Басова и др. // Лесное хозяйство. – 1999. - №4. – С. 51-52.
- Горбачев, В.Н. Почвообразование на карбонатных породах в подзоне сосновых лесов Средней Сибири / В.Н. Горбачев // Почвы сосновых лесов Сибири. – Красноярск: Институт леса и древесины им. В.Н. Сукачева, 1986. – С. 15–23.
- Емшанов, Д.Г. Методы пространственной экологии в изучении лесных экосистем / Д.Г. Емшанов. – Киев, 1999. – 220 с.
- Жуков, А.М. Грибные болезни лесов Верхнего Приобья / А.М. Жуков. – Новосибирск: Наука, 1978. – 242 с.
- Звягинцев, В.Б. Распространенность, вредоносность грибов комплекса Armillaria в лесах Беларуси и обоснование лесозащитных мероприятий: автореф. дис. на соиск. уч. степени канд. биол. наук: 06.01.11 / В.Б. Звягинцев. – Минская область, п. Прилуки, 2003. – 19с.
- Иванов, А.И. Агариковые грибы-ксилотрофы Пензенской области / А.И. Иванов // Микология и фитопатология. – 1981. - Том 15. - №3. – С. 192-197.
- Матусевич, Л.С. Лесопатологическое состояние еловых лесов на территории европейской части России / Л.С. Матусевич // Лесное хозяйство. - 2003. - №1. - С. 29.
- Наставление по отводу и таксации лесосек в лесах Российской Федерации. – М., 1993. – 72 с.
- Негруцкий, С.Ф. Корневая губка. - 2-е изд., перераб. и доп. / С.Ф. Негруцкий. – М.: Агропромиздат, 1986. - 196 с.
- Павлов, И.Н. Морфологические признаки грибов комплекса Armillaria mellea sencu lato циркумбореальной области / И.Н. Павлов, А.Г. Миронов, Н.П. Кутафьева // Хвойные бореальной зоны. – 2006. - №3. – С. 14-21.
- Петров, А.Н. Макромицеты пихтовых лесов Хамар-Дабана / А.Н. Петров // Лесопатологические исследования в Прибайкалье. – Иркутск: СИФИБР СО РАН СССР, 1989. - 148с.
- Погребняк, П.С. Общее лесоводство / П.С. Погребняк. – М.: Колос, 1968. - 440 с.
- Помазков, Ю. И. Иммунитет растений к болезням и вредителям: Учеб. пособие. / Гос. ком. СССР по нар. образованию. – М.: изд-во УДН, 1990. – 80 с.
- Почвы СССР / Афанасьева Т.В и др.; отв. ред. Г.В. Добровольский. – М.: Мысль, 1979. – 380 с.
- Романовский, М.Г. Продуктивность, устойчивость и биоразнообразие равнинных лесов европейской России / М.Г. Романовский. – М.: МГУЛ, 2002. – 97 с.
- Самойлов, Ю.И. Структура фитогенного поля на примере одиночных дубов Quercus robur (Fagaceae) / Ю.И. Самойлов // Ботанический журнал. – 1983. – Том 68. - №8. – С. 1022-1034.
- Санникова, Н.С. Микросистемный анализ ценопопуляций древесных растений / Н.С. Санникова. – Екатеринбург, 1992. – 56 с.
- Селочник, Н.Н. Распространенность и вредоносность опенка в дубравах Теллермановского леса / Н.Н. Селочник, Н.К. Кондрашова // Микология и фитопатология. – 1991. – Том 25. – Вып. 3 – С. 226-232.
- Селочник, Н.Н. Основные болезни дуба и их влияние на состояние дубрав Лесостепи / Н.Н. Селочник // Лесохоз. информация. – 1992. - №5. – С. 36-37.
- Слободян, Я.Н. Проблемы усыхания ельников Украинских Карпат / Я.Н. Слободян, Т.Г. Шпильчак, П.Я. Слободян и др. // / Ин-т леса НАН Беларуси: сб. науч. тр., 2001. - № 53. - С. 339-341.
- Соколов, Д.В. Корневая гниль от опенка и борьба с ней / Д.В. Соколов. – М.: Лесная промышленность, 1964. – 183 с.
- Стороженко, В.Г. Гнилевые фауты лесов Русской равнины / В.Г. Стороженко. – М., 2001. – 157 с.
- Федоров, Н.И. Корневые гнили хвойных пород / Н.И. Федоров. – М.: Лесная промышленность, 1984. – 160 с.
- Baumgartner, K., Distribution of Armillaria species in Califirnia / K. Baumgartner, D.M. Rizzo // Mycologia. – 2001. - vol. 93. – No.5. – P. 821-830.
- Bruhn, J.N. Distribution of Armillaria species in upland Ozark Mountain forests with respect to site, overstory species composition and oak decline / J.N. Bruhn, J.J. Wetteroff, J.D. Mihail et al. // Forest Pathol. - 2000. - 30, № 1. - P. 43-60.
- Hogg, E.H. Growth and dieback of aspen forests in northwestern Alberta, Canada, in relation to climate and insects / E.H. Hogg, J.P. Brandt, B. Kochtubajda // Can. J. Forest Res. - 2002. - 32, № 5. - P. 823-832.
- Korhonen, K. Fungi belonginc to the Genera Heterobasidion and Armilaria in Eurasia / K. Korhonen // Грибные сообщества лесных экосистем: материалы координационных исследований / Под. ред. В.Г. Стороженко, В.И. Крутова. – Москва-Петрозаводск, 2004. – Том 2. – С. 89-113.
- Korhonen, K. Interfertility and clonal size in the Armillaria mellea complex / K. Korhonen // Karstenia. – 1978. - №18. – P. 31-42.
- Robinson, R.M. Lesion formation and host response to infection by Armillaria ostoyae in the roots of western larch and Douglas-fir / R.M. Robinson, D.J. Morrison // Forest pathology. – 2001. - №31. – P. 371-385.
- Schwarze, F.W.M.R. Folge 8: Hallimasch-Arten / F.W.M.R. Schwarze, D. Ferner // AFZ/Wald. – 2003. – 58. - №14. – S. 718-719.
- Shaw III, C.G. Armillaria root disease. Agriculture handbook №691 / C.G. Shaw III, G.A. Kile. – Washington D.C. – 1991. – 231p.
- Vertui, F., Scots pine (Pinus sylvestris L.) die-back by unknown causes in the Aosta Valley, Italy / F. Vertui, F. Tagliaferro // Chemosphere. - 1998. - 36, № 4-5. - P. 1061-1065.
- Wahlstrom, K.T. Infection biology of armillaria species: in vitro pectinolytic activity, infection strategy, field distribution and host responses / K.T. Wahlstrom. – Uppsala: Swedish University of Agricultural Sciences, 1992. – 40 p.
|


