|
"Хвойные бореальной зоны" 2006г.,№1, с. 105-114
МЕЙОЗ ПРИ МИКРОСПОРОГЕНЕЗЕ У ПИХТЫ СИБИРСКОЙ
Бажина Е.В., Квитко О.В., Муратова Е.Н.
Институт леса им. В.Н.Сукачева СО РАН
Работа выполнена при финансовой поддержке ККФН-РФФИ, грант № 05-04-97717.
Исследовался мейоз у пихты сибирской (Abiessibirica Ledeb.) в различных экологических условиях: здоровые и усыхающие деревья, растущие в среднегорье Восточного Саяна (территория заповедника Столбы: долина р. Лалетина и верховья р. Калтат) и деревья, растущие в дендрарии Института леса им. В.Н. Сукачева СО РАН (окрестности г. Красноярск). Выявлены особенности мейоза у данного вида. У всех деревьев на разных стадиях обнаружены нарушения мейоза. Отмечены как общие, так и специфические типы нарушений. Предполагается, что некоторые мейотические нарушения могут привести к стерильности пыльцы и семян.
Meiosis of Siberian Fir (Abies sibirica Ledeb.) in the East Sayan middle mountains (territory of national park Stolby: basin of Laletina river and course of Kaltat river) and V. N. Sukachev Institute arboretum (Krasnoyarsk) has been studied. Microsporogenesis was investigated in details in healthy and drying trees. Features of A. sibirica meiosis were revealed. Common types of meiotic irregularities were observed in all studied trees. Meiotic irregularities were found in all trees at different stages. Specific irregularities were also revealed in some trees. It is proposed that some meiotic irregularities can be cause of pollen and seed sterility.
Введение
Пихта сибирская (Abiessibirica Ledeb.) является одним из основных лесообразующих сибирских видов. В горах Южной Сибири (Хамар-Дабан, Западный и Восточный Саяны) на пихтовые леса приходится около 16% лесопокрытой площади территории; по распространению она занимает третье место после кедра сибирского и лиственницы сибирской [1]. В последние десятилетия в горных экосистемах наблюдается усыхание пихтовых древостоев [2]. Усыхающие деревья характеризуются снижением жизнеспособности репродуктивных органов и нарушениями половой репродукции [3], что в перспективе ставит под угрозу существование пихтовых лесов. В связи с этим, становится актуальной не только организация мероприятий по охране растений в природе, но и разработка методов их сохранения в условиях культуры [4].
Одним из важнейших критических периодов полового размножения организмов является мейоз. Качество мужских гамет, а, следовательно, и успешность оплодотворения у растений зависит от прохождения мейоза, нарушения которого могут привести к стерильности пыльцы и семян. Мейоз у пихты сибирской детально не изучен. Краткое описание отдельных стадий приведено в монографии Т.П. Некрасовой и А.П. Рябинкова [5]. Некоторые нарушения в мейозе у пихты сибирской при промышленном загрязнении описаны Л.П. Преснухиной и Н.А. Калашник [6].
Цель настоящего исследования - изучение особенностей мейоза у пихты сибирской в различных экологических условиях среднегорья Восточного Саяна, а также при введении в культуру. При искусственном лесоразведении происходит адаптация организмов к новым условиям обитания. Анализ развития признаков и адаптивных реакций в культуре имеет большое значение для разработки общей теории онтогенеза и изучения механизмов реализации генетической программы онтогенетической адаптации. Исследование животных и растений в культуре показало, что новые условия существования часто являются для организма стрессовыми и разные организмы реагируют на них различным образом. При этом индивидуальная способность к адаптации достигается с помощью различных механизмов – физиологических, биохимических, генетических, цитогенетических.
Материал и методы
Исследования мейоза проводились в течение трех лет (2002-2004 гг.). Объектом исследований являлись деревья пихты сибирской, растущие в среднегорье Восточного Саяна (район заповедника Столбы): шесть - в долине р. Лалетина (высота 480-520 м над ур. моря) и шесть - в верховьях р. Калтат (высота 640-680 м над ур. моря), а также шесть деревьев, растущих в дендрарии Института леса им. В.Н. Сукачева. В верховьях р. Калтат были отобраны три усыхающих дерева пихты, а также три дерева без признаков усыхания (типичных). Характеристика деревьев представлена в таблице 1.
Дендрарий расположен в лесопарковой зоне Академгородка (г. Красноярск) на высокой (275 м над ур. м.) террасе левого берега Енисея на месте островного участка дерново-злаковой горной степи [7]. Климат региона резко континентальный, дендрарий относится к восточно-сибирскому варианту южно-таежной подзоны. Почва в дендрарии – дерново-карбонатная, характеризуется слабо-щелочной (близкой к нейтральной по рН солевому) реакцией среды, с невысоким содержанием гумуса и низкой степенью подвижности азота. Саженцы деревьев пихты привезены в дендрарий Института леса в 1977 г. из школьного отделения питомника НИИ плодоводства Сибири (г. Барнаул). Для выращивания посадочного материала были использованы семена местного (Алтай) происхождения [8]. По комплексу хозяйственно ценных признаков (габитус, семеношение, устойчивость к болезням и др.) отобранные деревья могут быть отнесены к категории лучших.
Материалом для исследования служили мужские генеративные почки отобранных деревьев. Материал выдерживали в фиксаторе 1-2 суток, затем переводили в 70%-й спирт на хранение. В качестве красителя использовали ацетогематоксилин. При проведении исследований было проанализировано 23208 микроспороцитов на разных стадиях мейоза и 28630 формирующихся пыльцевых зерен.
На каждой стадии мейоза учитывали общее количество мейоцитов в норме и с различного рода аномалиями в поведении хромосом. Исследования проводили с помощью микроскопа МБИ-6, фазы мейоза фотографировали на пленку "Микрат". Кроме того, часть препаратов анализировали под микроскопом Axioskop 20 (Карл Цейс, Германия) с видеокамерой и выходом на монитор «SONY Multiscan G200». Определяли также размеры и фертильность пыльцевых зерен. Статистическую обработку данных проводили по общепринятым методикам [9].
Результаты и обсуждение
Мейоз у пихты сибирской начинается и заканчивается весной. В третьей декаде апреля в микроспорангиях видны микроспороциты (материнские клетки пыльцы) округлой формы на стадии профазы первого деления. Диаметр их, как правило, в 1.3-1.5 раза больше, чем у окружающих клеток тапетума (рисунок 1а) и в среднем составляет 57.3±1.12 мкм (у клеток тапетума - 43.3±0.81 мкм).
У деревьев пихты сибирской можно выделить все стадии П I (лептотену, зиготену, пахитену, диплотену и диакинез). В мейоцитах насчитывается до 4 ядрышек (в дендрарии) и до 5 - (в естественных насаждениях); одно из них, как правило, крупнее и лучше прокрашено. Исследования показали, что ядрышки в мейоцитах пихты сибирской исчезают к концу диплотены, хотя ранее считалось, что они сохраняются до конца следующей стадии – диакинеза [5].
Мейоз у пихты сибирской проходит по классическому типу, с образованием 12 бивалентов, которые в основном правильно расходятся к противоположным полюсам. Морфология бивалентов определяется расположением и числом хиазм (рисунок 1б). У большинства исследованных деревьев наблюдалось до 3 хиазм на бивалент, лишь у одного дерева, растущего в дендрарии, насчитывалось до 5 хиазм. По литературным данным [10], у A. cephalonica и A. nordmanniana частота хиазм составляет в среднем 2.4 на бивалент.
Исследования показали, что развитие мужской генеративной сферы пихты сибирской в различных условиях произрастания имеет как общие, так и специфические особенности. Прежде всего, особенностью мейоза пихты, как и других хвойных видов, является его асинхронность. Асинхронное развитие мейоцитов характерно в целом для хвойных [5, 11-14], в том числе и для пихты сибирской. Однако если в естественных популяциях пихты в пределах одного микроспорангия встречаются микроспороциты на различных стадиях первого или второго деления мейоза [15], то в условиях дендрария наблюдались и клетки на стадии профазы первого деления и тетрады, и даже формирующиеся пыльцевые зерна (рисунок 1, в). При этом в микроспорангиях, расположенных в нижней части стробила, мейотическое деление идет быстрее по сравнению с микроспорангиями верхней части.
Общей особенностью мейоза пихты сибирской также является неодинаковая продолжительность разных фаз. Профаза первого деления является наиболее длительной стадией, она может продолжаться около недели.
Таблица 1 – Характеристика деревьев пихты сибирской
Место
произрастания |
Тип леса |
Возраст,
лет |
Высота,
м |
Диаметр на высоте
1.3 м, см |
Протяженность кроны, м |
Проекция кроны, м |
Долина
р. Лалетина |
Осинник разнотравный с примесью пихты |
110-130 |
11-14 |
20-26 |
9.0-12.0 |
5.0-6.0 |
Верховья
р. Калтат |
Пихтарник разнотравно-зеленомошно-вейниковый |
90-140 |
11-14 |
22-28 |
13.0-16.0 |
3.5-4.5 |
Дендрарий |
|
40-45 |
9-11 |
20-24 |
7-9 |
3.0-4.5 |
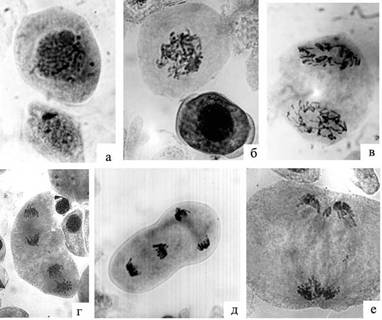
Рисунок 1 - Особенности развития мейоцитов и аномалий в дендрарии:
а – микроспороцит в профазе I и клетка тапетума; б – асинхронность развития микроспороцитов в условиях дендрария (профаза I и микроспора с формирующимися воздушными мешками; в – профаза второго деления; г – линейная ориентация веретен во втором деления мейоза (анафаза II); д – перпендикулярная ориентация веретен во втором делении мейоза (анафаза II); е – параллельная ориентация веретен во втором делении мейоза (анафаза II, слияние групп хромосом на одном полюсе).
Наиболее быстро проходят телофазы первого и второго делений, каждую из которых удалось наблюдать лишь у 0.05 % исследованных микроспороцитов (в естественных насаждениях) и у 0,2% (в дендрарии). Третьей особенностью пихты сибирской и, возможно, всего рода Abies [16], является четко идентифицируемая профаза второго деления (рисунок 1 в). У других видов хвойных она отсутствует и сразу начинается метафаза II, как, например, у сосны обыкновенной [14, 17].
Необходимо отметить особенности развития микроспороцитов в различных условиях. В естественных насаждениях мейоз начинался позднее (5-6 мая – в долине р. Лалетина и 10-13 мая – в верховьях р. Калтат), проходил очень быстрее и занимал не более 2 - 4-х суток, тогда как в условиях дендрария продолжительность его составляла в разные годы 18-21дн.. Сроки начала делений зависят от погодных условий года развития. Например, в 2002-2003 гг. в дендрарии первые мейотические деления были отмечены уже 24 апреля, а в 2004 г. - только 5 мая. Причиной такого различия в сроках начала делений, вероятно, является затяжная и холодная весна 2004 г. Постмейотическое развитие пыльцы у пихты сибирской в условиях дендрария в 2002-2003 гг. продолжалось до 11-13 мая, а в 2004 г. 13 мая еще шел мейоз.
Еще одной особенностью мейоза пихты в условиях дендрария является наличие наряду с нормальной ориентацией веретен на различных стадиях второго деления параллельная, линейная и перпендикулярная их ориентация (рисунок 1, г, д). Частота встречаемости мейоцитов с такими особенностями в разные годы составляла от 0,3 до 20,5% у разных деревьев. По мнению некоторых авторов [18, 19], в норме, во втором делении мейоза веретена должны быть ориентированы по отношению друг к другу под углом 60°. Параллельное расположение веретен может привести к слиянию групп хромосом на противоположных полюсах и к формированию пыльцевых зерен с диплоидным числом хромосом (рисунок 1, е). Однако процесс слияния параллельных веретен в значительной степени зависит от влияния факторов среды [20].
В большинстве изученных клеток мейоз проходил регулярно с правильным расхождением хромосом к противоположным полюсам. На разных стадиях мейоза были обнаружены и нарушения. Общее число клеток с нарушениями у деревьев пихты сибирской, растущих в долине р. Лалетина, составляло 3.0–4.3%, увеличиваясь в стрессовых условиях (в верховьях р. Калтат) до 7.5-9.6% у деревьев без признаков усыхания и 5.9-6.7% у усыхающих деревьев, а в дендрарии – до 7,0-20,4% (рисунок 2). Спектр нарушений у этих деревьев также значительно шире. Частота нарушений в мейозе на отдельных стадиях варьирует по годам и у разных деревьев. В первом делении мейоза у деревьев, растущих в долине р. Лалетина, наблюдались, в основном, нарушения общего типа, характерные для большинства исследованных деревьев.
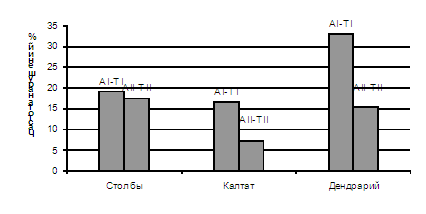
Рисунок 2 - Частота нарушений в мейозе у пихты сибирской.
К ним относятся: униваленты на стадии диакинеза, выброс хромосом в метафазе, хаотическое расхождение и отстающие хромосомы в анафазе, выброс хромосом и их фрагментов в телофазе.
У одного из исследованных деревьев (№ 6ст), наблюдались специфические типы нарушений: слипание отдельных хромосом или их крупных групп, что приводит к образованию поливалентных ассоциаций в метафазе первого деления. У этого дерева также была отмечена деспирализация и хаотическое расхождение групп хромосом к полюсам в анафазе. Мы полагаем, что нарушения в мейозе у дерева 6ст обусловлены генетически, вследствие его гибридной либо мутантной природы, однако, этот вопрос требует дальнейшего изучения. У деревьев, растущих в долине р. Калтат в анафазе наблюдались мосты и отстающие хромосомы. У усыхающих деревьев, кроме этого, было отмечено хаотическое расхождение в анафазе, выброс хромосом за пределы метафазной пластинки и 3-х полюсная А I (рисунок 3, а – ж)
Во втором делении мейоза у деревьев, растущих в долине р. Лалетина, среди общих нарушений наблюдали выброс хромосом в метафазе и слияние групп хромосом в анафазе. Специфический тип аномалий – фрагментация и слипание хромосом - был обнаружен у дерева № 22ст (рисунок 3, з,и). У деревьев № 6ст и 22ст отмечали также значительное количество микроспороцитов с линейной ориентацией веретен деления в анафазе второго деления и на стадии тетрад. Такая аномалия может привести к слиянию групп хромосом и формированию триад микроспор.
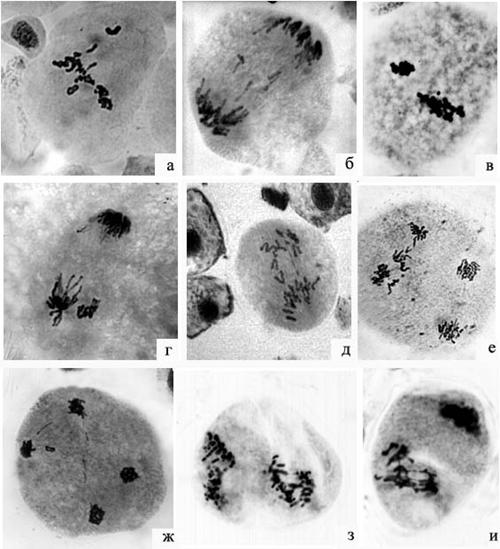
Рисунок 3 - Аномалии мейоза у деревьев пихты сибирской, растущих в экосистемах Восточного Саяна: а – выброс хромосом в метафазе I; б – отстающие хромосомы в анафазе I; в – поливалентные ассоциации в метафазе I; г - выброс группы хромосом в анафазе I; д –хаотическое расхождение и отстающие хромосомы в анафазе I; е – задержка группы хромосом на экваторе клетки в анафазе II; ж - выброс хромосом в анафазе II; з, и – фрагментация и слипание хромосом во втором делении мейоза
У деревьев, растущих в верховьях р. Калтат, в анафазе второго деления наблюдали мосты. У усыхающих деревьев обнаружены пятиполярные конфигурации в анафазе II, мосты, отстающие хромосомы, выбросы и хаотическое расхождение хромосом.
Исследования показали чрезвычайно широкий спектр нарушений мейоза у пихты в дендрарии. При этом в 2004 г. отмечена повышенная частота аномалий на разных стадиях мейоза для всех деревьев по сравнению с предыдущими годами исследования. Особенно велико было число нарушений в первом делении мейоза: от 6.8 до 42.8% у разных деревьев.
Отдельные типы нарушений (фрагментация хромосом в диакинезе, остаточное ядрышко на стадии прометафазы I и агглютинация хромосом в кольцо на стадии метафазы I, триполярные конфигурации и выброс групп хромосом на стадии ана-телофазы I, объединение ядер в диаде, «удлинение» хромосом, а также их хаотическое расположение на стадии метафазы II) отмечены только у деревьев, растущих в дендрарии и не встречались у деревьев, растущих в естественных экосистемах в окрестностях г. Красноярска (рисунок 4, а-д).
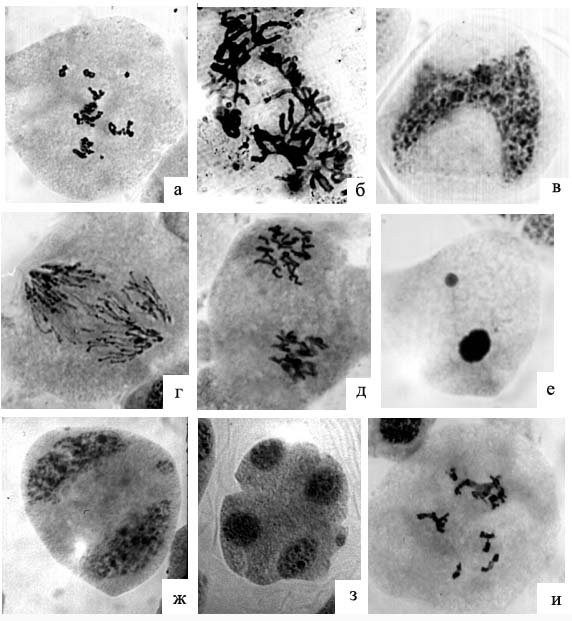
Рисунок 4 - Аномалии мейоза у пихты сибирской в культуре:
а – фрагментация хромосом в диакинезе; б – агглютинация хромосом в кольцо на стадии метафазы I; в – частичное слияние ядер в диаде; г – «удлинение» хромосом в анафазе I; е – агглютинация хроматина и выход ядрышка в цитоплазму клетки; ж – микроядро на стадии диады; з – микроядро на стадии тетрады; и – остаточное ядрышко на стадии диакинеза
Кроме того, у всех исследованных деревьев в этом году наблюдалась остановка развития микроспороцитов на стадии профазы первого деления. В таких клетках происходила агглютинация хроматина, часто наблюдался также выход ядрышка в цитоплазму клетки (рисунок 4, е). Впоследствии эти клетки, вероятно, дегенерировали. Количество микроспо-роцитов с данной аномалией составляло 0.20 – 2.21% у разных деревьев. Была отмечена специфичность развития некоторых аномалий в условиях дендрария. Так, в частности, наблюдалось обособление отстающих в анафазе хромосом в микроядра (рисунок 4, ж, з). Мосты в анафазе первого деления в некоторых случаях сохранялись и во втором делении мейоза, а также, вероятно, приводили к полному или частичному слиянию групп хромосом (рисунок 4 и).
У всех деревьев, растущих в дендрарии, наблюдались фрагментация хромосом, выбросы за пределы метафазной пластинки, хаотическое расхождение хромосом к полюсам и отстающие хромосомы, мосты, в том числе и множественные, триполярные конфигурации в анафазе второго деления. Частота этих аномалий у отдельных деревьев варьировала в разные годы от 0,1 до 25,6% от общего числа проанализированных на данной стадии мейоцитов.
К специфическим типам нарушений, отмеченным лишь у отдельных деревьев, относились поливалентные ассоциации в диакинезе, агглютинация хромосом в кольцо в метафазе первого деления, удлинение хромосом на стадии анафазы, хаотическое расположение хромосом в метафазе второго деления, микроядра в диаде и тетраде. Частота встречаемости таких нарушений значительно варьировала у разных деревьев. Особенно выделялось дерево № 1, у которого до 16,6 % мейоцитов имели специфические аномалии развития на различных стадиях деления. Еще у одного из исследованных деревьев (№ 6) в некоторых микроспороцитах наблюдалось большое количество хиазм в бивалентах (до 5), а у 8.5 % клеток на стадии диакинеза сохранялось ядрышко, которое имело неправильную форму (рисунок 4к).
Были также отмечены множественные нарушения (в 0,5-3,6 % клеток): мост + выброс хромосом за пределы метафазной пластинки, множественные мосты, отстающие хромосомы + выброс хромосом, отстающие хромосомы + мост на стадии ана- телофазы первого деления, мост + хаотическое расположение хромосом, хаотическое расхождение хромосом + мост, отстающие хромосомы + выброс на стадии ана- телофазы второго деления.
Еще один интересный и крайне редкий тип аномалий - асинхронность деления в пределах одного мейоцита: например, на одном полюсе второго мейотического деления может наблюдаться профаза II, на другом - метафаза II или в анафаза II. Отмечены также следующие сочетания: анафаза I - метафаза II, метафаза II - анафаза II, диада - анафаза II, телофаза II – тетрада, анафаза II- тетрада.
Аномалии в мейозе, как правило, элиминируются на стадии интеркинеза (диады, тетрады) и практически не влияют на жизнеспособность пыльцы. Однако исследования показали, что у всех исследованных деревьев наряду с нормальными тетрадами наблюдались триады и пентады микроспор. У усыхающих деревьев были отмечены также гексады микроспор, однако частота встречаемости подобной аномалии крайне низкая и составляет 0.6 и 0.7 % у разных деревьев.При микроспорогенезе наряду с нормальными микроспорами с 2 воздушными мешками были обнаружены микроспоры с 1 и 3 воздушными мешками, однако, частота таких аномалий была достаточно низкой и не превышала 2.04 %.
Морфология мейоза определяется взаимодействием конкретного генотипа, структуры хромосом и среды [21]. Особенная чувствительность к неблагоприятным условиям характерна для ювенильной стадии онтогенеза и начала репродуктивной фазы, когда процессы споро- и гаметогенеза входят в соответствие с температурным и световым режимом [22, 23]. Очевидно, увеличение продолжительности мейоза у деревьев пихты сибирской, наблюдающееся в дендрарии, является реакцией на экстремальные и непривычные условия среды. Аналогичное явление наблюдалось у сосны обыкновенной, произрастающей в стрессовых условиях [12, 19]. Вследствие увеличения продолжительности мейоза возрастает вероятность воздействия на клетки неблагоприятных факторов, что увеличивает количество аномальных клеток. Известно, что большинство стадий мейоза у хвойных очень чувствительны к изменениям температуры и другим неблагоприятным факторам среды [11, 24, 25- 31]. В частности, у пихты сибирской в условиях промышленного загрязнения уровень аномальных мейоцитов в 2,5-6,5 раз выше, чем в контрольных условиях, отмечается более широкий спектр нарушений структурного характера – фрагментаций, хромосомных мостов, кольцевых хромосом, выбросов и слипаний хромосом [6].
Сохранение хвойных растений в условиях культуры является одним из способов поддержания их генофонда. В то же время при адаптации организма к новым условиям произрастания может измениться его метаболизм и функционирование и, в целом, ход его онтогенеза. Исследования показали, что в условиях культуры частота аномалий у пихты выше, а спектр нарушений мейоза значительно шире по сравнению с естественными популяциями; наряду с аномалиями общего типа были отмечены специфические и множественные нарушения. Известно, что аномалии могут быть обусловлены мутациями генов, поскольку весь процесс мейоза находится под генетическим контролем [21, 32-34] или хромосомными перестройками [35]. Можно предположить, что более широкий спектр аномалий у деревьев пихты сибирской, растущих в дендрарии обусловлен и генетическими причинами, т.е. их гибридной или мутантной природой. Наличие хромосомных мутаций у пихты сибирской, в том числе и кольцевых хромосом, ранее было отмечено при изучении кариотипа [36]. Аномалии в поведении хромосом в мейозе, как правило, элиминируются еще до стадии тетрад и практически не влияют на качество образующихся пыльцевых зерен. Однако, некоторые нарушения (например, агглютинация хроматина) могут отражаться в неоднородности пыльцы по размеру и форме пыльцевых зерен и вызывать снижение ее фертильности [37]. И действительно, исследования показали наличие наряду с нормальными пыльцевыми зернами мелких и с аномальным числом воздушных мешков. Пыльца деревьев, растущих в естественных популяциях, как правило, обладает высокой фертильностью. Вероятно, отмеченные нарушения в микроспорогенезе могут объяснить снижение фертильности пыльцы у деревьев пихты сибирской, растущих в дендрарии (до 8-28% у разных деревьев).
Заключение
Таким образом, цитологическое изучение пихты сибирской выявило, что в целом, мейоз проходит по классическому типу. Некоторые типы нарушений отрицательно влияют на качество пыльцы, что может привести к затруднению процессов естественного возобновления данного вида. Выявленные особенности мейоза, наблюдаемые в стрессовых условиях (большая продолжительность и увеличение частоты аномалий) могут свидетельствовать об адаптации организмов.
Цитологическое изучение пыльцы пихты сибирской выявило, что в целом, мейоз проходит по классическому типу. Отмечены особенности мейоза у данного вида (асинхронность развития в пределах одного микроспорангия и микроспороцита, различную продолжительность фаз мейоза, нарушение расхождения хромосом). Анализ микроспорогенеза у пихты сибирской в условиях культуры выявил некоторые особенности мейоза: растянутость во времени и более широкий спектр аномалий по сравнению с деревьями пихты из естественных популяций. Большая продолжительность и повышенное число аномалий мейоза у деревьев, растущих в верховьях р. Калтат и в культуре, вероятно, обусловлены их реакцией на стрессовые условия. Некоторые аномалии мейоза могут приводить к снижению жизнеспособности пыльцы данного вида.
Библиографический список
1. Поликарпов, Н. П., Чебакова, Н. М., Назимова, Д. И. Климат и горные леса Южной Сибири.- Новосибирск: Наука, 1986. - 226 с.
2. Бажина, Е.В., Третьякова, И.Н // К проблеме усыхания пихтовых лесов. - Успехи современной биологии. – 2001.- Т. 121. - № 6. - С. 626-631.
3. Tretyakova, I.N. Bazhina, E.V. Structure of crown as well as pollen and seed viability of fir (Abies sibiica Ledeb.) in disturbed forest ecosystems of the Khamar-Daban Mts // Ecology (Bratislava). – 2000. -V 19. - № 3. - Р. 280-294.
4. Плотникова, Л. С. Научные основы интродукции и охраны культурных растений флоры СССР. - М.: Наука, 1988. - 264 с.
5. Некрасова, Т. П., Рябинков, А. П. Плодоношение пихты сибирской. - Новосибирск: Наука, 1978. - 150 с.
6. Преснухина, Л. Н., Калашник, Н. А. Микроспорогенез пихты сибирской (Abies sibirica Ledeb.) в условиях промышленного загрязнения // Актуальные проблемы генетики: Матер. II конф. Моск. об-ва генетиков и селекционеров им. Н. И. Вавилова, посвященной 115-летию со дня рождения акад. Н. И. Вавилова. - Москва, 20-21 февраля 2003г. - М., 2003. - Т. 2. - С. 321.
7. Мамаев, С. А., Дорофеева, Л.М., Александрова М.С., Звиргзд А.В., Лоскутов Р.И., Морякина, В.А., Петухова, И.П., Таран, И.В., Шкутко, Н.В., Фурст, Г.Г. Адаптация и изменчивость древесных растений в лесной зоне Евразии. - Екатеринбург: УИФ «Наука», 1993. - 136 с.
8. Лоскутов, Р.И. Декоративные древесные растения для озеленения городов и поселков.- Красноярск: изд-во Краснояр. ун-та, 1993. - 184 с.
9. Рокицкий, П.Ф. Биологическая статистика. - Минск: Высш. шк., 1973. – 320 с.
10. Sax, K., Sax, H. J. Chromosome number and morphology in the conifer // J. Arnold Arboretum.- 1933. - V. 14. - № 4. - P. 356-374.
11. Козубов, Г.М. Биология плодоношения хвойных на Севере. - Л.: Наука, 1974. - 136 с.
12. Рождественский, Ю. Ф. Особенности микроспорогенеза сосны обыкновенной на Урале и его зависимость от экологических факторов // Экология. - 1974. - № 1. - С. 49-53.
13. Мурая, Л. С., Буторина, А. К., Дудецкая Е. М. Мейоз при микроспорогенезе и развитие мужского гаметофита у лжетсуги в условиях интродукции // Лесоведение. - 1988. - № 5. - С. 37-44.
14. Муратова, Е. Н. Особенности мейоза сосны обыкновенной около северной границы ее ареала // Онтогенез. - 1995. - Т. 26. - № 2. - С. 158-169.
15. Бажина, Е.В., Квитко, О.В., Муратова, Е.Н. Цитогенетическое изучение пихты сибирской (Abies sibirica Ledeb.) в среднегорье Восточного Саяна // Актуальные проблемы генетики. Актуальные проблемы генетики. М-лы II-й конф. Моск. общ-ва генет. и селекц. им. Н. И. Вавилова, посвященной 115-летию со дня рождения акад. Н. И. Вавилова. – М, 2003. - Т. 2. - С. 262-264.
16. Mergen, F., Lester, D. T. Microsporogenesis in Abies // Silvae Genet. - 1961. - Bd 10. - Hf. 5. – S. 146-156.
17. Буторина, А. К., Высоцкий, А. А., Мурая, Л. С. Сравнительный цитогенети-ческий анализ форм сосны обыкновенной разной смолопродуктивности // Генетика. - 1982. - Т. 18. - № 12. - С. 2019-2029.
18. Mok, D. W. S., Peloquin, S. J. Three mechanisms of 2n pollen formation in diploid potatoes // Canad. J. Genet. and Cytol. - 1975. - Bd 17. - Hf. 2. - S. 217-225.
19. Буторина, А.К., Мурая, Л.С., Исаков Ю.Н.Характер прохождения мейоза при микроспорогенезе у сосны обыкновенной на суходоле и болоте // Половое размножение хвойных растений. - Новосибирск, 1985. - Тез. докл. II Всесоюзн. симп. - С. 7-10.
20. Veileux, R.E., Lauer, F.I. Variation of 2n pollen production in clones of Solanum phureja Juz. and Buk. – Theor. Appl. Genet. - 1981. - V. 59. - P. 95.
21. Соснихина, С.П., Федотова, Ю.С., Смирнов В.Г., Михайлова Е.И., Богданов Ю.Ф. Изучение генетического контроля мейоза у ржи // Генетика. - 1994. - Т.30. - № 8. - С. 1043-1056.
22. Шкутко, Н.В. Развитие генеративных почек у хвойных растений, интродуцированных в БССР// Половая репродукция хвойных. - Новосибирск: Наука. 1973. - Т.2. - С.132-134.
23. Гаврилов, И.А., Буторина, А.К. Цитогенетика тсуги канадской в условиях интродукции в Воронежской области // Лесоведение. - 2005. - № 3. - С. 60-65.
24. Круклис, М. В. Мейоз и формирование пыльцы у лиственницы Чекановского (Larix czekanowskii Sz.) // Изменчивость древесных растений Сибири. - Красноярск: Институт леса и древесины АН СССР, 1974. - С. 20-34.
25. Яковлев, А. В. О влиянии низких температур на микроспорогенез сосны обыкновенной // Лесоведение. - 1978. - № 6. - С. 51-55.
26. Christiansen, H. On the effect of low temperature on meiosis and pollen fertility in Larix decidua Mill. // Silvae Genet. - 1960. - B 9. - H. 3. - S. 72-78.
27. Kantor, J., Chira, E. Microsporogenesis in some species of Abies // Sbornic Vysoke Skoly Zemedelske Brne (Rada C). - 1965. - № 3. - P. 179-185.
28. Chandler, C., Mavrodineanu S. Meiosis in Larix laricina Koch // Contrib. Boyce Thompson Inst. - 1965. - V. 23. - № 4. - P. 67-76.
29. Eriksson, G. Temperature response of pollen mother cells in Larix and its importance for pollen formation // Stud. Forest. Suec. - 1968. - № 63. - 131 p.
30. Ekberg, I., Eriksson, G., Jonsson, A. Meiosis in pollen mother cells of Pinus contorta // Hereditas. -1972. - B 71. - H. 2. - S. 313-324.
31. Luomajoki, A. Effects of temperature on spermatophyte male meiosis // Hereditas. - 1977. - V. 85. - P. 33-48.
32. Голубовская, И.Н. Генетический контроль поведения хромосом в мейозе// Цитология и генетика мейоза. - М.: Наука, 1975. - С. 127-139.
33. Голубовская, И. Н. Экспериментальное исследование генного контроля мейоза у кукурузы // Теоретические основы селекции. Новосибирск: Наука, 1985. - С. 119-135.
34. Соснихина, С.П., Смирнов, В.Г., Михайлова, Е.И., Егорова, Л.Ф. Нарушение гомологичного синапсиса у мейотических мутантов диплоидной ржи // Генетика. - 1994. - Т. 30. - № 4. - С. 488-494.
35. Хвостова, В.В., Ячевская, Г.Л. Перестройки хромосом в мейозе// Цитология и генетика мейоза. - М.: Наука, 1975. - С. 232-262.
36. Муратова, Е. Н., Матвеева, М.В. Кариологические особенности пихты сибирской в различных условиях произрастания // Экология. - 1996. - № 2. - С. 96-102.
37. Пожидаева, И.М., Буторина, А.К.., Исаков, Ю.Н. Нарушения в мейозе при микроспорогенезе и уровень фертильности у сосны обыкновенной // Половое размножение хвойных растений: Тез. докл. II Всесоюз. симпоз. - Новосибирск, 10-12 сент. 1985г., - Новосибирск, 1985. - С. 31.
|


