|
"Хвойные бореальной зоны" 2004г. с.128-140
ГЕНОМНЫЕ И ХРОМОСОМНЫЕ МУТАЦИИ У СОСНЫ ОБЫКНОВЕННОЙ (PINUSSYLVESTRISL.) В ЭКСТРЕМАЛЬНЫХ УСЛОВИЯХ ПРОИЗРАСТАНИЯ
Муратова Е.Н., Седельникова Т.С
Институт леса им. В. Н. Сукачева СО РАН, Красноярск, Россия
Работа выполнена при частичной финансовой поддержке Красноярского краевого фонда науки (12F0006C), интеграционного проекта СО РАН № 145 и проекта РАН 12.1.
Представлены материалы по цитогенетическому изучению популяций сосны обыкновенной, произрастающих в экстремальных условиях Южного Забайкалья (южная граница видового ареала), Центральной Якутии (северная граница видового ареала), Красноярского края (радиоактивное и техногенное загрязнение), Томской области (болота) и Волгоградской области (за пределами южной границы ареала вида). Выявлено значительное количество геномных и хромосомных нарушений. Проведен сравнительный анализ спектра и встречаемости мутаций в изученных насаждениях сосны с соответствующими показателями для популяций этого вида из других экстремальных условий произрастания.
The data on cytogenetic studies of Scots pine populations growing under extreme conditions of South Zabaikalje (the southern boundary of species area), Central Yakutia (the northern boundary of species area), Krasnoyarsk region (radioactive and industrial pollution), Tomsk region (swamps), and Volgograd region (outside the southern boundary of species area) are presented. A great number of genome and chromosome abnormalities were revealed. A comparative analysis of spectrum and frequency of mutations in studied populations of pine and ones growing under other extreme conditions was performed.
Введение
Сосна обыкновенная (PinussylvestrisL.) является важнейшим лесообразователем бореальной зоны Евразии. Данный вид имеет обширный ареал и распространен на территории Евразии в разнообразных физико-географических и экологических условиях [1-2]. У сосны обыкновенной выделено пять подвидов, множество разновидностей и форм. Полиморфизм популяций по многим признакам свидетельствует о наличии у данного вида широкой нормы реакции. Важное значение имеет выявление адаптивных особенностей данного вида в экстремальных условиях среды, формирующих популяции с повышенным уровнем мутабильности и своеобразием генотипической структуры.
Кариологические и цитогенетические
исследования популяций сосны обыкновенной в экстремальных условиях среды проводились в различных регионах [3-25]. В кариотипе этого вида выявлен широкий спектр хромосомных аномалии, которые встречаются в экстремальных условиях произрастания с повышенной частотой.
В настоящем сообщении обобщены материалы по изучению хромосомных аномалий в разных частях ареала вида: около южной границы ареала (в Южном Забайкалье и в окрестностях г. Читы); около северной границы ареала (в Туруханском районе Красноярского края и в Центральной Якутии); на болотах Западной Сибири (Томская обл.), в том числе у деревьев с "ведьминой метлой" и на территориях, подвергшихся влиянию осушительной мелиорации; за пределами видового ареала (сухая степь юга России, Волгоградская область); в условиях антропогенного стресса (Красноярская ТЭЦ, Железногорск, Горнохимический комбинат). Собственные данные сравниваются с результатами других авторов, изучавших этот вид по единой методике.
Материал и методы
Объектами настоящих исследований послужила сосна обыкновенная (PinussylvestrisL.) из различных мест произрастания. Кариологический анализ производился по общепринятой для хвойных методике [26] с собственными модификациями. Кончики корешков проростков семян обрабатывали 0,5 % раствором колхицина в течение 6-8 ч, фиксировали спиртов о-уксусной смесью (3:1), окрашивали ацетогематоксилином и готовили давленые препараты по стандартным методикам.
Хромосомные нарушения анализировали на стадии метафазы традиционным методом с применением колхицина. Кроме того, нарушения митоза изучали на стадиях ана- и телофазы [27]. Учитывали общее количество просмотренных клеток, находящихся на разных стадиях митоза, и количество патологических митозов. В изученных клетках определяли суммарное количество перестроек, затем производили пересчет встречаемости мутаций на 100 клеток.
Результаты
В кариотипе сосны обыкновенной имеется 24 хромосомы (2n=24); данный вид является диплоидом с основным числом х=12; длина одной хромосомы составляет 10-18 мкм. Десять пар хромосом (1-Х) являются метацентрическими. Две наиболее короткие хромосомы XI и XII пар в разных наборах могут быть как метацентрическими, так и субметацентрическими.
Сотрудниками лаборатории лесной генетики и селекции Института леса проведены комплексные исследования сосны обыкновенной в Южном Забайкалье [4], у южной границы ареала вида. Найдены деревья различных форм, с отклонениями роста и развития, в том числе, возможно, мутантного происхождения. Изучение кариотипа и особенностей клеточного деления показало большое число геномных и хромосомных нарушений, а также нарушений митоза. Например, в семенном потомстве деревьев с наростами (или опухолями) встречались гипо - и гиперанеушгоиды (с 2n=22 и 28). У многих растений обнаружена миксоплоидия, когда наряду с диплоидным числом хромосом (2n=2х=24) встречались тетраплоидное (2n=4х=48) и более высокой степени плоидности). Миксоплоидия отмечена не только в семенном потомстве, но у материнских растений, в меристематической зоне оснований хвои. Это деревья с наростами, полукарлики и некоторые типичные [6].
По характеру хромосомных нарушений можно отметить кольцевые хромосомы разных типов, ди- и трицентрические хромосомы. Наблюдались типичные кольцевые хромосомы, дицентрические и двойные кольцевые хромосомы, ацентрические кольца и кольца, надетые на обычную палочкообразную хромосому, кольцевые хромосомы с фрагментами и без фрагментов. Иногда встречались отдельные фрагменты. Кольцевые хромосомы встречались чаще, чем хромосомы с несколькими центромерами. Эти хромосомы не являются добавочными, а соответствуют нормальным хромосомам диплоидного набора.
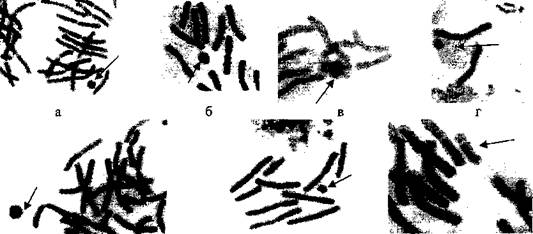
При наличии одной или нескольких кольцевых хромосом число хромосом не изменялось. Если в клетке имелась дицентрическая хромосома, число хромосом уменьшалось до 2n=23. Хромосомные аномалии имелись только в отдельных клетках. Геномные и хромосомные нарушения у сосны обыкновенной в Южном Забайкалье представлены на рисунке 1.
Изучение митоза у этих же растений показало, что размах изменчивости по числу поврежденных ана - и телофаз составляет от 0,2 до 8,0%. Встречались следующие аномалии: одиночные и парные мосты, одиночные, парные и множественные фрагменты, отставание целых хромосом или их фрагментов, хаотическое расхождение хромосом.
Основную массу аберраций составляли одиночные фрагменты и мосты без фрагментов. Клетки с мостами и фрагментами встречались редко.
Повышенная частота патологических митозов отмечена у деревьев с опухолями и полукарликов.
 

ж
Рисунок 1 - Геномные и хромосомные нарушения у сосны обыкновенной в Южном Забайкалье: а -клетка с тетраплоидным набором хромосом (2n=4х=48); б - метафазная пластинка с кольцевой хромосомой; в, г - фрагменты метафазных пластинок с кольцевой хромосомой; д - надетая кольцевая хромосома; е, ж - дицентрическая хромосома; з - ди- и трицентическая хромосомы; и -дицентрическая хромосома и парные фрагменты. Хромосомные нарушения показаны стрелками. Окраска ацетогематоксилином. Об. 90х, ок. 10
Высокая частота возникновения хромосомных аномалий у сосны у южной границы ареала, вероятно, связана с воздействием неблагоприятных факторов в период развития растений [28; 4; 6]. Нарушения митоза у сосны обыкновенной в Южном Забайкалье представлены на рисунке 2.
Изучение хромосом сосны обыкновенной из Центральной Якутии показало, что северные популяции отличаются от центральных и южных увеличением длины хромосом, повышенным числом вторичных перетяжек и особенностями их локализации [16-17]. В потомстве деревьев с нарушением габитуса (полукарлики, деревья с опухолями, с плакучей формой кроны) наблюдалась миксоплоидия: большинство клеток обладало диплоидным числом хромосом, но среди них были триплоидные (2n=3х~36) и тетраплоидные (2n=4х=48).
При цитогенетическом изучении Р. sylvestrisиз Центральной Якутии выявлены также хромосомные мутации. Некоторые из них сходны с хромосомными нарушениями, которые были обнаружены в Центральной Тыве [28] и в Южном Забайкалье [6]. Среди нарушений отмечены: мелкие и крупные ацентрические кольца, кольцевые хромосомы (одна или две), кольца с фрагментами и без фрагментов, кольцевая хромосома 8-образной конфигурации, надетые кольцевые хромосомы, ди- и трицентрические хромосомы, дицентрики вместе с кольцевыми хромосомами, одиночные и парные фрагменты и другие аномалии. Кроме того, в популяции Центральной Якутии отмечен редкий тип аномалий: в некоторых случаях кариотип содержал одну асимметричную хромосому (субакро- или акроцентрическую), в то время как второй гомолог был метацентриком.
Рисунок 2 - Нарушения митоза у сосны обыкновенной в Южном Забайкалье: а, б - одиночный фрагмент в анафазе; в - парный фрагмент в анафазе; г, д - парные фрагменты в метафазе; е -хаотическое расхождение хромосом в анафазе; ж - кольцевая и отстающая хромосомы в анафазе; з - отстающие фрагменты; и, к - отстающие хромосомы; л - разорванный одиночный мост; м, н -разорванный парный мост; о - разорванный парный мост и фрагменты. Окраска ацетогематоксилином. Об. 90х, ок. 10
Присутствие асимметричной хромосомы с очень коротким плечом в кариотипе Р. syhestrisсвидетельствует о делеции в гетерозиготном состоянии. Хромосомные нарушения у сосны обыкновенной в Центральной Якутии представлены на рисунке 3. Частота аномалий в Центральной Якутии низкая - от 0,2 до 2,7%. Это значительно ниже, чем частота хромосомных мутаций у сосны обыкновенной в Южном Забайкалье.
Только у четырех растений отмечено более 1% аномалий. Это типичное дерево, дерево с опухолями и два дерева с плакучей формой кроны. Аномалии митоза встречались с частотой от 1,6 до 4,0%. Среди них обнаружены одиночные и парные фрагменты, хромосомные и хроматидные мосты с фрагментами и без них, отстающие и забегающие вперед хромосомы, хаотическое расхождение хромосом.
Д еж
Рисунок 3 - Хромосомные нарушения у сосны обыкновенной в Центральной Якутии: а, б -кольцевая хромосома; в - надетая кольцевая хромосома; г, д - ацентрическое кольцо; е -фрагмент; ж - хромосома с делецией. Хромосомные нарушения показаны стрелками. Окраска ацетогематоксилином. Об. 90х, ок. 10
В анафазе была обнаружена кольцевая хромосома 8-образной конфигурации. Анализ мейоза при микроспорогенезе у сосны обыкновенной в Центральной Якутии выявил аномалии в развитии генеративной сферы. На разных стадиях были обнаружены мейотические нарушения. Наблюдались следующие типы аномалий: униваленты на стадии профазы I, отстающие хромосомы, аномалии веретена на стадии метафазы I, мосты, фрагменты, мосты с фрагментами на стадии анафазы I, трехполюсные митозы, мосты и фрагметы на стадиях анафазы II и телофазы II, триады, стерильная пыльца и другие аномалии. Часть аномалий может быть связана с действием неблагоприятных факторов среды, а другие могут быть вызваны хромосомными перестройками или мейотическими мутациями [7; 16-17]. Нарушения мейоза у сосны обыкновенной в Центральной Якутии представлены на рисунке 4. Проведено изучение митозов у сосны обыкновенной в районе Горно химического комбината (окр. г.Железногорска): в зоне радиационного загрязнения (вариант 1 - в районе первого отстойника, вариант 2 - около полигона, по дороге на пос. Балчуг) и за пределами 30- км-й зоны (контроль). В зоне загрязнения процент нарушений митоза в анателофазе был значительно выше, чем в контроле - соответственно, 18,1%, 18,4% и - 5,3% - от числа изученных клеток [25]. У сосны обыкновенной в районе промышленных выбросов Красноярской ТЭЦ также выявлены хромосомные нарушения, однако их частота не выше, чем в контроле [5]. Кариологическое исследование популяций сосны обыкновенной на болотах (экстремальные условия) и суходолах (типичные условия) Западной Сибири показало, что на болотах увеличиваются число вторичных перетяжек и частота их встречаемости; выявлены хромосомы с четырьмя вторичными перетяжками [8-10; 23]. Кроме того, кариотипы болотных популяций характеризуются более диффузным распределением вторич-ных перетяжек на плечах ядрышкообразующих хромосом.
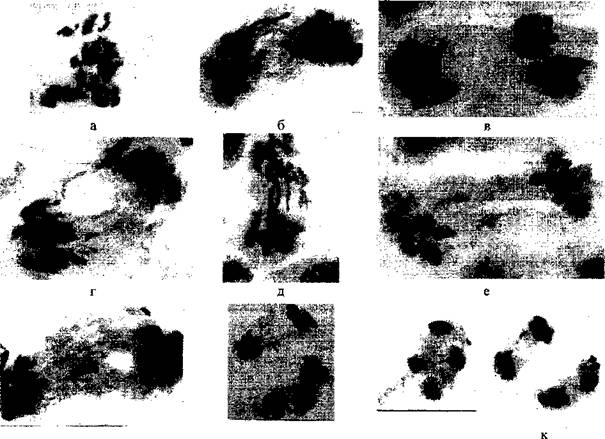
У сосны обыкновенной на болотах выявлен широкий спектр геномных и хромосомных мутаций, а также нарушения митозов. Геномные мутации выражались в наличии хромосомных мозаи и миксоплоидов (вместе с клетками, имеющими нормальное число хромосом, имелись полиплоидные). Среди структурных мутаций чаще всего наблюдались аберрации преимущественно типа кольцевых структур (кольца, кольцевые хромосомы, иногда с фрагментами). В единичных случаях наблюдались дицентрики. Отдельные метафазы содержали кольца, надетые на обычную хромосому. Частота хромосомных перестроек в кариотипах сосны верхового болота и суходола составляет 0,4%, в кариотипе сосны низинного болота - 1,3%. Примеры геномных и хромосомных мутаций у болотной формы сосны обыкновенной представлены на рисунке 5.
У сосны обыкновенной с "ведьминой метлой" число геномных и хромосомных мутаций выше, чем у нормальных деревьев на болоте [10]. Деревья с «ведьминой метлой» в отдельных случаях содержали триплоидные и тетраплоидные клетки. В митозе отмечены отстающие и хаотически расходящиеся хромосомы, мосты, с-митоз. Сосна с "ведьминой метлой" отличается еще большей активностью ядрышкообразующих зон, выражающейся в увеличении числа
вторичных перетяжек до 6 на хромосому, а также нарушением структуры и функций ядрышек в интерфазных ядрах [11]. Были обнаружены аномальные формы ядрышек и так называемое «остаточное ядрышко» в метафазе митоза (рисунок 5в). Подобные аномалии описаны у древесных растений в зоне Чернобыля [14]. Они могут возникать под действием каких-либо стрессовых факторов.
ж з и
Рисунок 4 - Нарушения мейоза у сосны обыкновенной в Центральной Якутии: а - десинапсис в метафазе I; б — отстающие фрагменты хромосом в анафазе I; в - расщепление веретена в анафазе 1; г, д, е - мосты и разрывы мостов в анафазе I; ж - фрагмент в анафазе I; з - фрагмент в анафазе II; и - мост в анафазе II; к - разорванный мост в анафазе II. Об. 90х, ок. 1
 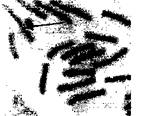 
в б а
Рисунок 5 – Геномные и хромосомные мутации в болотных популяциях сосны обыкновенной Томской области: а – клетка с тетраплоидным набором хромосом (2п=4х=48); б – метафазная пластинка с фрагментом; в – аномальное ядрышко в интерфазном ядре у сосны с «ведьминой метлой». Хромосомный фрагмент и аномальное ядрышко показаны стрелкой. Окраска ацетогематоксилином. Об. 90х, ок. 10
В целом, частота встречаемости аберраций и нуклеолярная активность хромосом на болотах выше по сравнению с суходолами, что может быть связано как с экологическими факторами, так и с генотипической структурой популяций изученных видов. На осушенных территориях повышенная частота хромосомных мутаций, возможно, связана с нарушением экологической обстановки после проведения мелиоративных работ. В связи с этим сбор семян для лесокультурной практики с таких деревьев нежелателен. Выявленные особенности свидетельствуют о биологической пластичности сосны обыкновенной в гидроморфных условиях произрастания и могут служить индикаторами реакции растений на различные стрессовые факторы.
Таблица 1 – Встречаемость хромосомных нарушений у сосны обыкновенной у южной и северной границ ареала
Вариант |
Анализ метафаз |
Анализ анателофаз |
изучено проростков |
изучено клеток |
число /
процент
аномалий |
изучено проростков |
изучено клеток |
число /
процент
аномалий |
Южное Забайкалье |
К-18 |
33 |
621 |
66/10.7 |
45 |
263 |
24/ 8,0 |
К-35 |
29 |
286 |
26/8.7 |
50 |
441 |
23/ 4,6 |
К-33-1 |
32 |
392 |
9/2.2 |
48 |
314 |
5/ 1,7 |
101 |
38 |
-604 |
5/0.8 |
48 |
665 |
4/0,6 |
55 |
44 |
856 |
1/0.1 |
41 |
591 |
3/0,5 |
1 |
51 |
666 |
14/2.0 |
36 |
400 |
9/2,2 |
К-31 |
34 |
577 |
- |
32 |
428 |
1/0,2 |
К-157 |
28 |
521 |
4/08 |
40 |
673 |
13/1,9 |
Центральная Якутия |
2т |
53 |
650 |
10/1.5 |
24 |
437 |
7/1,6 |
1оп |
48 |
975 |
6/0.6 |
25 |
428 |
15/3,5 |
1п |
51 |
596 |
16/2.7 |
37 |
547 |
15/2,7 |
Зп |
50 |
1269 |
14/1.1 |
26 |
475 |
19/4,0 |
Южный предел сплошного распространения сосны обыкновенной проходит по границе лесостепной и степной зон, где ареал вида разрывается. Здесь сосна произрастает в виде изолированных, или островных, боров, что является особенностью ее современного ареала [1]. В более южных районах России сосна обыкновенная в составе естественных насаждений не встречается. Были исследованы геномные и хромосомные мутации в потомстве интродуцированных популяций сосны обыкновенной за пределами южной границы ее ареала в условиях сухой степи (Волгоградская область). Изученные деревья в искусственных посадках сосны характеризовались проявлениями вторичного роста и нарушениями габитуса.
Большая часть клеток в кариотипе сосны из Волгоградской области содержит нормальное число хромосом 2n=24. Однако в семенном потомстве некоторых растении отмечена миксоплоидность: вместе с нормальными клетками встречались три- и тетрагоюидные. Среди общего числа изученных метафаз клетки с геномными нарушениями составляют 1,4%. Структурные перестройки хромосом выявлены в 13% изученных клеток (таблица 2). По характеру нарушений наиболее часто встречались кольцевые структуры. В отдельных случаях в клетках изученных растений обнаружены дицентрики, иногда фрагменты. Частота встречаемости их намного ниже, чем кольцевых хромосом. Дицентрическая хромосома по своим параметрам отличается от остальных хромосом набора: она примерно в полтора раза длиннее и содержит дополнительную центромеру. Примеры хромосомных аномалий у сосны обыкновенной за пределами ареала приведены на рисунке 5.
Таблица 2 – Встречаемость и спектр геномных и хромосомных нарушений в кариотипе сосны обыкновенной в экстремальных условиях сухой степи юга России
Типы перестроек |
Число метафаз с перестройкой, шт. |
Встречаемость
метафаз с перестройкой, % |
Кольцевые хромосомы с фрагментами или без фрагментов |
10 |
6,9 |
Надетые кольцевые хромосомы |
1 |
0,7 |
Ацентрические кольца |
1 |
0,7 |
Фрагменты |
4 |
2,7 |
Дицентрические хромосомы |
3 |
2,0 |
Геномные нарушения |
2 |
1,4 |
Широкий спектр и повышенная встречаемость хромосомных мутаций у сосны обыкновенной за пределами ареала в условиях сухой степи могут быть связаны с экстремально высокими температурами и низким количеством осадков в течение вегетационного периода, нехарактерными для типичных местопроизрастаний данного вида.
Обсуждение
Для сравнения вместе с собственными результатами в таблице 3 приводятся данные других авторов по встречаемости хромосомных и геномных нарушений в популяциях сосны обыкновенной из экстремальных условий произрастания с различными лимитирующими факторами. Анализ показывает, что возникновение геномных мутаций, связанных с изменением числа хромосом (миксоплоидия, неуплоидия), характерно для маргинальных популяций сосны возле северной и южной границ ареала, а также в пессимальных экологических условиях. У деревьев с нарушениями габитуса, признаками замедленного роста (карликовые формы) и отклонениями в развитии (опухолевидные образования, «ведьмина метла») встречаемость геномных нарушений выше, чем у типичных деревьев [6; 4; 10; 23; 16-17]. Гаплоидные растения, а также миксоплоиды обнаружены у сеянцев сосны в теплицах [3]. При антропогенном стрессе (промышленные и радиоактивные загрязнения) геномные нарушения как реакция на ухудшение условий произрастания для насаждений сосны не отмечены. Появление анеуплоидов, миксоплоидов и полиплоидов в популяциях сосны обыкновенной не является исключительным событием. Полиплоидия представляет собой генетический механизм, связанный с изменением дозы гена. Благодаря полиплоидии на базе исходного генома возникают новые адаптивные формы, имеющие морфологические, анатомические и биохимические отличия от него. Эволюционное преимущество полиплоидов связано с появлением и реализацией новых путей онтогенетического и популяционного гомеостаза. Образование полиплоидных клеток в насаждениях сосны обыкновенной за пределами естественного ареала вида может рассматриваться как один из регуляторных механизмов, позволяющих поддерживать определенный уровень устойчивости популяции в неблагоприятных условиях среды.
Появление хромосомных перестроек в метафазных и анателофазных клетках в популяциях сосны обыкновенной на северной (Якутия} и южной (Тыва, Южное Забайкалье) границах ареала и за его пределами (Волгоградская область), а также в зоне экологического пессимума на болотах (Западная Сибирь) и в горах (Южный Урал) в условиях изоляции также связывается с воздействием различных неблагоприятных факторов [13; 4; 8-10; 23; 16-17]. Спектры хромосомных аберраций в популяциях сосны из условий произрастания с экстремальными природными факторами сходны. Возникновение мутаций в этих случаях может иметь адаптивное значение.Наиболее часто встречающиеся в метафазных клетках сосны кольцевые структуры возникают при делециях обоих плеч хромосомы с образованием среднего фрагмента с двумя «липкими» концами и двух концевых фрагментов. Концевые фрагменты объединяются, а средний замыкается в кольцо.
Таблица 3 - Встречаемость геномных и хромосомных мутаций у сосны обыкновенной в экстремальных условиях произрастания
Происхождение и год сбора материала |
Действующий фактор |
Частота
геном-ных
нару-шений,
% |
Частота хромосомных нарушений, % |
Ссылки |
мета-фазы |
ана-
тело-фазы |
Окрестности г. Чернобыля, 1987-1992, 1986-1998 гг. |
Радиоактивное загрязнение |
- |
11.0 |
0.1-2.3 1.5-8.0 |
14; 24 |
Словакия, искусственные насаждения, 1997 г. |
Загрязнение тяжелыми металлами |
- |
1.0-1.3 |
- |
15 |
Челябинская обл., окр. г. Сатки и г. Карабаша, 1992-93 гг. |
Техногенное загрязнение |
- |
15.0-
32.1 |
17.4-20.2 |
13; 20;
22 |
Окр. г. Красноярска, 1986 г. |
Выбросы ТЭЦ |
- |
0.1-07 |
1.0-1.9 |
5 |
Окр. г. Железногорска Красноярского края, 1998 г. |
Техногенное и радиоактивное загрязнение |
- |
- |
18.1-18.4 |
25 |
Воронежская обл., окр. с. Истобного |
Радиоактивное загрязнение, электромагнитное излучение |
- |
- |
4.1 |
19 |
Окрестности г. Екатеринбурга» |
Техногенное загрязнение |
- |
- |
6.6-72.9 |
18 |
Брянская обл., окрестности
г. Новозыбковска,
1991-1993 гг. |
Низкие дозы радиации, выбросы транспорта |
- |
- |
7.0 |
33 |
Украина, деревья с «ведьминой метлой» |
Обработка семян супермутагенами (НДММ) |
- |
- |
5.7-9.4 |
38 |
Челябинская обл., Кировская обл. |
Теплицы с полиэтиленовым покрытием |
0.00140.00 28 |
- |
- |
3 |
Томская обл., олиготрофное
болото, типичные деревья,
1986 г. |
Переувлажнение, низкая трофность почвы |
- |
0.6 |
- |
8; 9 |
Томская обл., олиготрофное
болото, деревья с «ведьминой метлой», 1998 г. |
Тоже |
0.92-10.1 |
10.5 |
5.3 |
10; 11;
23 |
Томская обл., евтрофное болото, 1986 г. |
Мелиоративные работы |
- |
1.5 |
- |
12 |
Горы Южного Урала |
Изоляция |
- |
- |
3.6-11.9 |
13 |
Центральная Якутия, деревья
с нарушениями габитуса,
1988 |
Северная граница ареала |
Есть |
0.2-2.7 |
1.6-4.0 |
16; 17 |
Южное Забайкалье, типичные деревья, 1978-87 гг. |
Южная граница ареала |
0.6-4.3 |
0.2-10.3 |
0.5-2.2 |
4; 6 |
Южное Забайкалье, деревья с
нарушениями габитуса и
отклонениями в развитии,
1978-87 гг. |
Южная граница ареала |
0,1-8.5 |
0.1-15.7 |
0.2-8.0 |
4; 6 |
Центральная Тыва, типичные деревья, 1978-85 гг. |
Южная граница ареала |
4.0-26.0 |
- |
2.0-7.7 |
4 |
Центральная Тыва, деревья с
признаками замедленного
роста, 1978-85 гг. |
Южная граница ареала |
7.0-51.2 |
- |
3.8-12.6 |
4 |
Волгоградская обл., деревья с нарушениями габитуса, 1998 г. |
За пределами ареала, сухая степь |
1.4 |
13.0 |
- |
39; 40 |
Рисунок 6 - Хромосомные мутации (отмечены стрелкой) у сосны обыкновенной из Волгоградской области: а, б - ацентрическое кольцо; в - надетая кольцевая хромосома; г - дицентрическая хромосома. Ацетогематоксилин. Об. 90х. Ок. 10х
Кольцевые хромосомы растений являются нестабильными структурами – они изменяются в числе и размерах, а также теряются в процессе размножения клеток. Мутационная форма надетости кольца на палочковидную хромосому наблюдается при соматическом кроссинговере. Образование дицентрика свидетельствует о наличии асимметрич-ной транслокации в клетке. В этом случае происходит перераспределение генетического материала между хромосомами, приводящее к дополнительному резерву изменчивости, увеличению диапазона нормы реакции, перестройке регуляторной системы генома. Участие транслокаций в дивергенции популяций может являться эволюционным фактором [29]. Основные типы анателофазных патологий включают нарушения структурно- морфологической целостности хромосомы или общей картины митоза. Присутствие в анафазе одиночных фрагментов свидетельствует о делециях хроматидного типа, парных - хромосомного. Мосты с фрагментами образуются при асимметричных транслокациях. Одиночные мосты также считаются хроматидными, а парные - хромосомными [27]. Парные мосты представляют собой дицентрические хромосомы. Во время митоза они образуют мост, растягиваясь между дочерними хромосомами. В телофазе мосты рвутся, но могут сохраняться 12-15 циклов [30].
Хромосомные и хроматидные мосты, фрагменты, относятся к патологиям митоза, связанным с нарушениями синтеза ДНК и РНК, разрывом молекулы ДНК. Другие патологии митоза (отставание, неправильное расхождение, выбросы за пределы метафазной пластинки) могут быть результатом аномалии центромерного участка или нарушения функции сократительных белков ахроматинового веретена [31]. Представляет интерес и образование "остаточных ядрышек", формирование которых может быть связано с ингибированием синтеза белка. Функционирование ядрышка в течение всего митотического цикла рассматривается как компенсаторный механизм клетки при супрессии генов в результате действия стрессовых факторов [14]. С-митоз является одной из форм патологии митоза, дезорганизующей митотический аппарат при воздействии статмокинетических ядов [32]. Он встречается при интоксикации и различных патологических процессах в клетке.Согласно полученным результатам и данным, приведенным в таблице 3, высокий процент хромосомных мутаций в митозе отмечается в популяциях сосны, испытывающих техногенную или радиационную нагрузку. В этих случаях, помимо таких «типичных» нарушений, как фрагменты, мосты, отстающие и забегающие хромосомы [20; 22; 15], возникают специфические повреждения «жесткого» типа - агглютинация хромосом, вакуолизация цитоплазмы, кариомиксис, а также амитоз, микроядра [33; 14; 18]. Нарушения жесткого типа, как правило, являются летальными для клеток и приводят к угнетению и гибели растения. Характерной особенностью кариотипов хвойных из экстремальных условий является увеличение числа и частоты встречаемости вторичных перетяжек (нуклеолярных локусов) в хромосомах. Вторичные перетяжки рассматриваются как дополнительные маркеры, позволяющие идентифицировать отдельные пары хромосом, и являются важными в функциональном отношении структурами. Установлено, что в области вторичной перетяжки расположены ядрышковые организаторы, ответственные за синтез
рибосомной РНК [34-35]. Рибосомальные цистроны в составе полигенного комплекса контролируют ростовые процессы у растений [36]. Широкое распространение нуклеолярного полиморфизма у хвойных имеет эволюционное значение, поскольку активность рибосомных генов связана с адаптацией деревьев к определенным факторам окружающей среды. Возникновение добавочных вторичных перетяжек в хромосомах растений может быть связано как с включением дополнительных ядрышковых организаторов, так и со структурными перестройками этих районов (транслокациями), что свидетельствует о неодинаковой транскрипционной активности ядрышкообразующих хромосом. Диффузный характер локализации вторичных перетяжек может быть обусловлен присутствием перестроек типа инверсий [36-37]. Таким образом, в потомстве деревьев из экстремальных местопроизрастаний выявлена высокая частота встречаемости геномных и хромосомных мутаций и изменчивость нуклеолярных локусов. Появление мутантных форм свидетельствует о сдвигах в системе физиологического гомеостаза, дифферен-цировке популяций, перестройке их генотипов соответственно изменению условий среды. Наряду с высокой элиминацией их носителей, хромосомные и генные мутации приводят к формированию адаптированного генома, повышают устойчивость популяций в экстремальных условиях произрастания, а также играют большую роль в процессах микроэволюции и видообразования.
Библиографический список
- Правдин Л.Ф. Сосна обыкновенная.- М.: Наука, 1964.- 190 с.
- Бобров Е. Г. Лесообразующие хвойные СССР. - Л.: Наука. Ленингр. отд- ние, 1978.-189 с.
- Исаков Ю. Н., Буторина А. К., Мурая Л. С. Обнаружение спонтанных гаплоидов у сосны обыкновенной и перспективы их использования в лесной генетике и селекции // Генетика. - 1981. - Т. 17, №
4. - С. 701-707. 4.Сосна обыкновенная в Южной Сибири. - Красноярск, 1988. - 150 с.
- Буторина А. К. Факторы эволюции кариотипов древесных // Успехи современной биологии. - 1989. - Т. 108, вып. 3 (6). - С. 342-357. 7.
- Муратова Е.Н., Зубарева О.Н. Цитогенетическая характеристика сосны обыкновенной [PinussylvestrisL.) в районе выбросов тепловой электростанции // Изв. СО АН СССР. Сер. биол. наук.- 1990. № 3.-С. 36-41.
- Муратова Е.Н. Хромосомные мутации у сосны обыкновенной в Южном Забайкалье // Изв. АН СССР. Сер. биол. 1991. № 5. С. 689-699.
- Муратова Е. Н. Особенности мейоза сосны обыкновенной (PinussylvestrisL.) около северной границы ее ареала // Онтогенез. - 1995. - Т. 26, № 2. - С. 158-169.
- Седельникова Т. С, Муратова Е. Н. Генеративные органы и кариотип сосны обыкновенной на олиготрофных болотах Западной Сибири // Лесоведение. - 1991. - № 3.-С. 34-44.
- Седельникова Т.С, Муратова Е.Н. Кариологическое исследование болотной формы сосны обыкновенной // Сибирский биол. журн. -1992. - Вып. 5. - С. 50-61.
- Седельникова Т. С, Муратова Е. Н. Цитологическое изучение сосны обыкновенной типа "ведьмина метла" на болоте // Цитология. - 1999. - Т. 41, № 12. - С. 1082.
- Седельникова Т. С, Муратова Е. Н. Кариологическое изучение Pinussylvestris(Pinaceae) с "ведьминой метлой", растущей на болоте // Ботан. ж. - 2001. - Т. 86, N 12. - С. 50-60.
- Муратова Е.Н., Седельникова Т.С. Кариологическое исследование болотных и суходольных популяций сосны обыкновенной (Pinus sylvestris L.) // Экология.- 1993.- № 6.- С. 41-50.
- Калашник Н.А., Шафикова Л.М. Биоиндикация в лесных экосистемах и городах, подвергшихся промышленному загрязнению. Цитогенетический метод биоиндикации // Биоценотическая характеристика хвойных лесов и мониторинг лесных экосистем Башкортостана.- Уфа: Гилем, 1998.- С. 255-260.
- Butorina А. К., Kosichenko N. Е., Isakov Yu. N., Pozhidaeva I. M. The effect of irradiation from the Chernobyl nuclear power plant accident on the cytogenetic behavior and anatomy of trees // Cytogenetics study of forest trees and shrub species: Proc. First IUFRO Cytogenet. Work. Party S2.04-08 Symp. Zagreb, 1997.-P. 211-226.
- Micieta K., Murin G. The use of Pinus sylvestris L. and Pinus nigra Arnold as bioindicator species for environmental pollution // Cytogenetic Studies of forest trees and shrub species. Proc. First IUFRO cytogenetics Working Party S2.04-08 Symp. Zagreb, 1997. P. 253-263.
- Muratova E. N. Cytological and karyological studies on Scots pine (Pinus sylvestris L.) near the northern border of its area // Генетика и селекция - на службе лесу. Матер, межд. научно-практ. конф. - Воронеж, 1997а,- С. 33-44.
- Muratova E. N. Cytogenetical study on Scots pine {Pinus sylvestris L.) in the Central
Yakutia // Cytogenetic studies of forest trees and shrub species: Proc. First IUFRO Cytogenetics
Working Party 2.04-08 Symp. Zagreb, 1997b. -
P. 157-177.
- Zuyeva G. V., Babushkina L. G., Makhneva S. G. Chromosomal anomalies in Pinus sylvestris seeds germinated under polluted air conditions // Cytogenetic Studies of forest trees and shrub species. Proc. First IUFRO cytogenetics Working Party S2.04-08 Symp. Zagreb, 1997.-P. 285-291.
- Миронов А. Н., Панкратьева Е. A. Изменчивость цитологических показателей у сосны обыкновенной (PinussylvestrisL.) под влиянием мутагенов среды // Цитология. -1999.-Т. 41, №12. -С. 1070-1071.
- Шафикова Л. М. Цитогенетические особенности сосны обыкновенной в условиях промышленного загрязнения: Автореф. канд. дис. биол. н. - Красноярск, 1999. - 20 с.
- Цитогенетическая характеристика семенного потомства некоторых видов древесных растений в условиях антропогенного загрязнения г. Воронежа / А.К. Буторина, А.Н. Калаев, Т.А. Вострикова, О.Е. Мягкова // Цитология. - 2000. - Т. 42, №2.-С. 196-201.
- Шафикова Л. М., Калашник Н. А. Характеристика кариотипа сосны обыкновенной из условий промышленного загрязнения // Лесоведение. - 2000. - № 2. -С. 30-36.
- Седельникова Т. С, Муратова Е. Н., Ефремов С. П. «Ведьмина метла» у сосны обыкновенной на болоте. Морфологические и цитологические особенности // Лесоведение. - 2000. - № 6. - С. 78-81.
- Кальченко В. А., Федотов И.С. Генетические эффекты острого и хронического воздействия ионизи-рующих излучений на Pinus sylvestris L., произрастающих в зоне отчуждения
Чернобыльской АЭС // Генетика.- 2001.- Т. V, № 4.- С. 437-447.
- Патология митоза у хвойных как показатель нарушенности гомеостаза / Е.Н.Муратова, Т.С. Седельникова,
О.С.Владимирова, Т.В. Карпюк, Л.В.Дмитриева // Гомеостаз лесных экосистем. Материалы Х-го Межд. симп. «Концепция гомеостаза: теоретические, экспериментальные и прикладные аспекты».-Новосибирск: Наука, 2001.-С. 113-119.27. Методика кариологического изучения хвойных пород / Л.Ф. Правдин, В.А. Бударагин, М.В. Круклис, О.П. Шершукова // Лесоведение, 1972. - N 2. - С. 67-75.
- Бочков Н.П., Демин Ю.С., Лучник Н.В. Классификация и методы учета хромосомных аберраций в соматических клетках // Генетика.- 1972.- Т. 8.- N 5.- С. 133-141.
- Сунцов А.В. Спонтанные хромосомные мутации у сосны обыкновенной в Центральной Туве // Изв. СО АН СССР. Сер. биол. наук. - 1982. - Вып. 3,- N15.-С. 55-58.
- Дубинин Н.П. Общая генетика. - М.: Наука, 1986.- 559 с.
- Зосимович В.П., Кунах В.А. Уровень, типы и происхождение аберраций хромосом в культуре изолированных тканей растений // Генетика. - 1975. - Т. 11, N 6. - С. 37-46.
- Алов И. А. Цитофизиология и патология митоза. - М.: Медицина, 1972. - 263 с.
- Hsu Т. С, Humphrey R. M., Somers С. Е. Persistent nucleoli in animal cells following treatments with fluorodeoxyuridine and thymidine // Exp. Cell Research. - 1964. - N 33. - P. 74-77.
- Butorina A.K., Evstratov N. The first detected case of amitosis in pine // Forest genetics. 1996. V. 3. N. 3. P. 137-136.
- Гриф В.Г., Агапова Н.Д. К методике описания кариотипов растений // Ботан. журн. - 1986. - Т. 71, N 4. - С. 550-553.
- Кикнадзе И.И. Функциональная организация хромосом. - Л.: Наука. Ленингр. отд-е, 1972.-211с.
- Bianche G., Bassi P., Carmona M.J., Maggini F. Contenuto di DNA ribosomale e regioni organizzatrici nucleolari in ornithogalum montanum (Liliaceae) // Atti. Assoc, genet, ital. -1980.-vol. 25.-P. 44-45.
- Буторина А.К. Факторы эволюции кариотипов древесных // Успехи соврем, биологии.- 1989.- Т. 108.- Вып. 3 (6).- С. 342-357.
- Nicoloff H., Anastassova-Kristeva M., Kunzell G. Changes in nucleolar organizer activity due to segmental interchange between satellite chromosomes in barley // Biol. Zentrbl. -1977.-vol. 96.-P. 223-227.
- Молотков П.И., Кириченко О.И., Бенгус Ю.В. О происхождении «ведьминой метлы» у сосны обыкновенной // Цитология и генетика.- 1989. - Т. 23.- N 4.- С. 14-19.
- Sedelnikova T.S. Genome and chromosome mutations of Pinus sylvestris L. outside the southern timberline of the species range // Cytogenetic studies of forest trees and shrubs - Review, Present status, and look on the Future: Abstr. 2 IUFRO Cytogenetic Working Party S 12.04-08 Symp. Graz, 1998. P. 28.
- Седельникова Т.С. Хромосомные и геномные мутации у сосны обыкновенной в Нижнем Поволжье // Лесоведение. - 2003. -N6.-С. 28-33.
|






