|
"Хвойные бореальной зоны" 2003г.,№1, с.
МИКРОЭВОЛЮЦИОННЫЕ АСПЕКТЫ ВНУТРИВИДОВОЙ ДИФФЕРЕНЦИАЦИИ ЛИСТВЕННИЦЫ СУКАЧЕВА НА УРАЛЕ
Путенихин В.П.
Ботанический сад-институт УНЦ РАН, Республика Башкортостан, Уфа.
Формирование дизъюнктивной популяционной системы лиственницы Сукачева на Южном-Северном Урале связано со сложными процессами миграции и взаимодействия хвойных видов в послеледниковое время, а также с явлениями инбридинга и генетического дрейфа на внутрипопуляционном уровне. Микроэволюционное развитие крайне северных популяций протекает на фоне интрогрессивной гибридизации с лиственницей сибирской. Повышенный полиморфизм этих, а также высокогорных популяций, по всей видимости, поддерживается естественным отбором. обеспечивая их адаптивность и благоприятные эволюционные перспективы. В целом, относительно низкая внутрипопуляционная изменчивость лиственницы Сукачева (в основном, в южной половине Урала) компенсируется достаточно высокой степенью межпопуляционной дифференциации.
Establishing the disjunctive population system of the Urals' Sukachev Larch is effected by complex processes of migration and interrelations of conifer species during the post-Ice Age, and the inbreeding and genetic drift phenomena at the intra-species level. Micro-evolutionary development of the most northern populations correlates with introgressive hybridization of Sukachev and Siberian Larch. Excessive polymorphism of these - as well as mountain - populations is probably supported by natural selection that ensures their adaptability and good evolutionary prospects. In general, relatively low intra-population Sukachev Larch changeability (typical for the Southern Urals) is balanced by a high degree of inter-population differentiation.
Лиственница Сукачева, внутривидовая сибирской (Larixsibirica Ledeb.). дифференциация, популяция, интрогрессивная В настоящем сообщении анализируются гибридизация, микроэволюция. процессы микроэволюционного развития
Ранее нами [26, 27] на основе многомерного популяций лиственницы Сукачева на Урале. анализа комплекса морфологических признаков Формирование популяций в генеративных органов была изучена историческом аспекте. внутривидовая дифференциация лиственницы Имеющиеся материалы по истории Сукачева (Larixsukaczewii Dyl.) на Урале и расселения лиственницы показывают, что в выделены 11 локальных фенотипически и плейстоцене, в периоды ледниковийгенотипически различающихся популяций: межледниковий, лиственница на Урале и маргинальная уральская (крайне южная), сопредельных территориях, то сокращая свои центральная и высокогорная южноуральские, площади (при этом отступая в на юг), то башкирская предуральская, пермско-камская, захватывая новые местообитания, являлась среднеуральская, центральная и высокогорная характерным компонентом перигляциальной североуральские, приполярно-уральская, печоро-растительности [3, 14]. Начиная со второй тиманская, полярно-уральская. Было установлено, половины голоцена происходили редукция что уровень внутрипопуляционного разнообразия лиственничных лесов и формирование нарастает в северном направлении и с современной колониальной структуры [2, 3, 11, продвижением в высокогорья. В формировании 21, 30, 35], однако на Приполярном и Полярном фенотипической структуры крайне северных Урале положение лиственницы в растительном популяций выявлена важная роль ландшафте оставалось гораздо более устойчивым интрогрессивной гибридизации с лиственницей [8,11].
Дифференциация популяций лиственницы Сукачева в регионе в связи с историческими процессами может рисоваться следующим образом. Полное пространственное отделение лиственницы в Башкирском Предуралье, где она и до этого произрастала лишь спорадически [23, 24, 33], возможно, произошло во время массированного расширения площадей ели и широколиственных лесов в конце бореального начале атлантического периода (9-7 тысяч лет тому назад). Более того [14, 24], рефугиум лиственницы Сукачева (и ели сибирской), возможно, оставался здесь со времен плейстоцена, обеспечивая таким образом гораздо более длительную изоляцию башкирскопредуральской популяции.
Вторичное вторжение темнохвойных пород в горную область Южного Урала в начале субатлантической эпохи и еще более раннее расширение зоны обитания сосны и березы [21, 24, 30], привели к дальнейшей дизъюнкции лиственницы и, возможно, с этим периодом (7,5-5 тысяч лет назад) связано обособление высокогорных лиственничных лесов Южного Урала (высокогорная южноуральская популяция), являющихся, таким образом, отголоском древнего «холодного» плейстоценового ландшафта. В дальнейшем лиственница на среднегорных хребтах и в Зауралье (центральная южноуральская популяция), по всей видимости, эволюционировала самостоятельно, все более и более уступая позиции сосне и березе. Последующая географическая (и популяционногенетическая) дивергенция популяций
лиственницы Сукачева, в частности, маргинальной уральской популяции на южном пределе ареала, обусловлена ксерофитизацией климата в последнем отрезке голоцена.
Итак, на Южном Урале пространственная, фенотипическая и генетическая дифференциация вида на несколько локальных популяций может объясняться сложными взаимоотношениями с другими хвойными и лиственными видами, а также значительным изменением природноклиматических условий в регионе в конце плейстоцена и в течение голоцена.
На Среднем и Северном Урале дополнительные причины, возможно, играли существенную роль. Так, среднеуральская популяция, главной примечательной чертой которой является ее «мелкошишечный фенотипический облик», вероятно, имеет тесную историческую и генетическую преемственность с холодной перигляциальной лиственничной лесотундрой (или сосново-лиственничноберезовой лесостепью), которая существовала у края плейстоценового ледника (как раз на Среднем Урале) [3, 14]. Возможно также, что после деградации ледника лиственница из южноуральских и южнее располагавшихся рефугиумов, мигрируя на север [11, 12, 35], более быстро продвигалась вдоль выровненного восточного и (или) западного макросклона; обогнув Средний Урал (и локализовав таким образом мелкошишечную популяцию) она достигла Северного Урала и сформировала здесь относительно крупношишечную популяцию.
Нельзя также исключить, что формирование пермско-камской предуральской популяции объясняется миграцией лиственницы Сукачева из ее плейстоценовых рефугиумов, располагавшихся на юге Русской равнины, где она отмечается на протяжении всего плейстоцена [7] - именно отсюда она распространялась на север европейской части России (и, вероятно, в Скандинавию) вдоль Волги, Камы и других водных артерий [12, 15, 38]. Соединение в районах Среднего, Северного Урала и Камского Предуралья популяций различного происхождения проясняет, с одной стороны, их выраженную фенотипическую дифференциацию, а с другой стороны - возникновение общей популяционной группы (отличной от южноуральской) благодаря взаимодействию и обмену генетическим материалом на протяжении почти всего голоцена. Выделение высокогорной североуральской популяции, без сомнения, произошло из состава центральной североуральской по окончании бореального периода, когда темнохвойная тайга (в том числе, кедровая) расчленила ареал лиственницы по высотной трансекте [4, 5, 11].
Важнейшей предпосылкой формирования приполярно-и полярно уральских популяций было, по всей видимости, продвижение лиственницы в новые суровые регионы (освободившиеся после отступления ледника) и процесс интрогрессивной гибридизации с лиственницей сибирской, обусловивший своеобразие микроэволюционных процессов и значительное возрастание внутрипспуляционной изменчивости. Формирование этих популяций, на наш взгляд, уже нельзя рассматривать в отрыве от процесса исторического движения лиственницы сибирской.
По окончании ледникового периода расселение лиственницы сибирской шло из Центральной Сибири и южно-сибирских горных систем в Западную Сибирь [1, 8, 22, 35], повидимому, вдоль Иртыша и Оби на север и северо-запад, поскольку на западе климатическая и гидрографическая обстановка (относительно быстрое потепление, обширные участки степей, а также болот на месте обширных подпрудноледниковых морей) была неблагоприятной для этого вида [8, 13]. Пути расселения древесных растений вдоль речных бассейнов, минуя водоразделы, считаются наиболее
предпочтительными как в холодные, так и в ксеротермические эпохи [18]. Аналогичным, например, было движение лиственницы Сукачева в европейской части России на север вдоль крупных рек [12].
В начале голоцена лиственница сибирская достигла Урала в его полярно-уральской части, конкретнее - в нижнем течение реки Обь [2, 35].
Примерно в то же время и лиственница Сукачева появилась на северном пределе Урала, двигаясь вслед за отступающим ледником. Эти виды встретились здесь, после чего и начался процесс их интрогрессивного смешения. В настоящее время, гибридизация охватила как Приполярный Урал, так и западную часть Западной Сибири [19, 27].
Дифференциация приполярной и полярной популяций друг от друга, по всей видимости дело недалекого исторического прошлого: оно связано как с определенным различием природноклиматических условий Приполярного и Полярного Урала, так и со степенью затронутости интрогрессией той и другой популяции, а также с влиянием южнее и западнее расположенных популяций (североуральских и печоротиманских). В отношении печоро-тиманской популяции наиболее вероятным предположением является ее происхождение от равнинных популяций лиственницы Сукачева, которые захватывали в голоцене северо-восток европейской части России, а на подходе к Уралу смешивались с двигающимися на север уральскими популяциями.
Для оценки среднего сходства изучаемых популяций с некоей точкой дивергенции был предложен так называемый метод сравнения с «прапопуляцией» [9, 10]. Полученные этим методом данные (таблица 1, рисунок 1), на наш взгляд, служат достаточно убедительным подкреплением изложенных выше соображений о путях миграции и взаимоотношениях уральских популяций лиственницы Сукачева.
Так, наибольшей близостью к «прародительской популяции» характеризуются центральные и высокогорные южно-и североуральские популяции. Отметим, что более крупные размеры генеративных органов в основных популяциях лиственницы Южного и Северного Урала по сравнению с другими частями ареала [8, 17, 25], также указывают, исходя из представлений В.Н. Сукачева [29] о крупношишечных и крупносемянных прародителях лиственниц, на большую древность этих популяций. Гораздо большая удаленность других популяций от «исходной» (по обобщенным расстояниям) может служить подтверждением иных путей их миграции (особенно, предуральских, см. выше), а также исторически позднего.
Таблица 1- Сравнение популяций лиственницы Сукачева с «прапопуляцией »
Популяция |
Обозначение |
D |
R |
Сумма рангов |
Место |
1 |
2 |
3 |
4 |
5 |
6 |
Маргинальная уральская Центральная южноуральская Высокогорная южноуральская Башкирская предуральская Среднеуральская Пермско-камская Центральная североуральская Высокогорная североуральская Приполярно-уральская Печоро-тиманская Полярно-уральская |
1-МР/УР 2-ЦТ/ЮЖ 3-ВС/ЮЖ 4-БШ/ПР 5-СР/УР 6-ПМ/КМ 7-ЦТ/СВ 8-ВС/СВ 9-ПР/УР 10-ПЧ/ТМ 11-ПЛ/УР |
2,28±0,344 (2) 1,96±0,238(1) 2,70±0,483 (5) 2,53±0,409 (4) 2,84±0,612(8) 2,94±0,606(11) 2(52±0,433 (3) 2,83±0,541 (7) 2,78±0,516(6) 2,87±0,562 (9) 2,92±0,502(10) |
0,974±0,0014(9) 0,979±0,0013(7) 0,982±0,0013(4) 0,969±0,0016 (10) 0,984±0,0011 (3) 0,980±0,0012(6) 0,989±0,0012(1) 0,988±0,0013(2) 0,977±0,0016(8) 0,981±0,0013 (5) 0,933±0,0083 (П) |
11 8 9 14 11 17 4 9 14 14 21 |
5-6 2 3-4 7-9 5-6 10 1 3-4 7-9 7-9 11 |
Примечание. D - расстояние Махаланобиса [20] от «прапопуляции» (по комплексу количественных признаков). R
- показатель сходства [9, 10] с «прапопуляцией» (по комплексу качественных признаков). В столбце 3 в скобках указаны порядковые места популяций после ранжирования D от наименьшего значения к наибольшему, в столбце 4 в скобках после ранжирования R от наибольшего значения к наименьшему. В столбце 5 "Сумма рангов" - сумма порядковых мест популяций из столбцов 3 и 4. В столбце 6 - итоговое порядковое место популяции (1 - наиболее близка к «прапопуляции», 11 - наиболее удалена от «прапопуляции». Различия популяций с «прапопуляцией» по всем
показателям D и R статистически значимы.
Микроэволюция вида на Урале.
На |
фоне |
проведенного |
исторического |
экскурса |
полученные |
ранее |
данные |
по |
внутривидовой |
структуре |
и |
изменчивости |
популяций [26, 27] позволяют высказать некоторые соображения эволюционного плана.
Историческая преемственность насаждений лиственницы из высокогорий Южного Урала с древними ранне-голоценовыми лесами позволяет поставить эту высокогорную популяцию на первое (фундаментальное) место в современной популяционной структуре вида в южной оконечности Урала. Ее относительно повышенное фенотипическое (и генетическое) формирования популяций Приполярного и Полярного Урала. Разнообразие показывает, что автохтонная высокогорная популяция до сих пор является носителем определенной части генетического потенциала, аккумулированного на предыдущих стадиях развития. Сходное объяснение высокого полиморфизма высказывалось и для высокогорных популяций ели Шренка (Piceaschrenkiana Fisch. et С. А. Меу) [31]. Этот внутрипопуляционный полиморфизм, который обычно поддерживается естественным отбором [32, 36], обеспечивает адаптивность лиственницы в условиях высокогорий и является, по нашему мнению, основой ее сохранения, а при благоприятных обстоятельствах - и расселения в регионе. Аналогичным образом можно определить и микроэволюционное развитие высокогорной североуральской популяции.
Значительное сокращение размеров других популяций и субпопуляций южной половины Урала, несомненно, приводит к возрастанию роли генетического дрейфа, что обусловливает снижение внутрипопуляционной изменчивости. Так, эволюция лиственницы на Уфимском плато (слабо-полиморфная башкирско-предуральская популяция) протекала в голоцене в условиях изоляции и жесткой конкуренции с темнохвойными породами. Учитывая давнее прекращение потока генов с Южного Урала, вероятное влияние таких факторов, как генетический дрейф и инбридинг, роль которых возрастает в изолятах [6, 28, 32], можно говорить о предопределенности ее эволюционной судьбы (полном вытеснении) в случае резкого изменения условий существования. Этот вывод согласуется с результатами дендроэкологического
прогнозирования выживаемости лиственничников на Уфимском плато [16].
Причиной наблюдаемого снижения внутрипопуляционной изменчивости в центральной южноуральской и маргинальной уральской популяциях может быть элиминация части генотипов на основе движущего отбора, протекающего в современных меняющихся условиях, а также в результате дрейфа генов [32], что, без сомнения, обуславливает дефицит фенотипов и высокий уровень инбридинга, выявленный также в результате генетического анализа [34].
У лиственницы на Среднем и Северном Урале (и в Печоро-Тиманском Предуралье), как было установлено, внутрипопуляционная изменчивость постепенно возрастает, однако, современное резкое дробление и сокращение ее здешнего ареала, локализация и изоляция субпопуляций, по-видимому, приводит в действие те же микроэволюционные факторы, что и на Южном Урале. Иная микроэволюционная обстановка прослеживается в обширных по площади приполярно-и полярно-уральских популяциях, занимающих разнообразные в экологическом отношении местообитания. Высокая внутрипопуляционная изменчивость свидетельствует о достаточной стабильности и адаптивности этих насаждений, что может являться следствием активно протекающего стабилизирующего отбора.
Не менее важным фактором, воздействующим здесь на микроэволюционные процессы, выступает интрогрессивная гибридизация. Исходя из характера движения лиственничных лесов в начале голоцена и климатического режима этого времени логично предположить, что первоначальное преимущество в расселении здесь приобрела лиственница сибирская - достигнув Полярного Урала, она стала распространяться и южнее по уральской горной системе, гибридизируя с лиственницей Сукачева. Отголоском такой экспансии, вероятно, и являются те промежуточные фенотипы, которые в небольшом количестве обнаруживаются в горах Северного и даже в высокогорьях Южного Урала [11, 26, 27]. В современную эпоху потепления процесс этот, очевидно, не только приостановился, но и приобрел обратное направление - теперь уже лиственница Сукачева на Полярном Урале интенсивно продвигается на восток -в пределы ареала лиственницы сибирской.
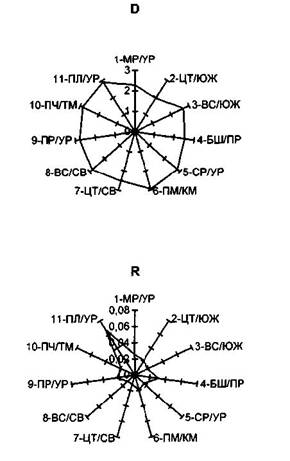
Рисунок 1- Сходство популяций лиственницы Сукачева с «прапопуляцией» (D - по расстоянию Махаланобиса, R - по показателю сходства Животовского). Названия популяций см. в таблице 1.
Внутривидовые микроэволюционные процессы, в частности, важная роль различных форм естественного отбора, дрейфа генов, изоляции и миграции в дифференциации популяций ранее были охарактеризованы при изучении лиственницы западной {Larixoccidentalis Nutt.) [37, 42] и американской {Larixlaricina (Du Roi) С. Koch) [39, 40, 41]. Выявленная у этих видов в отдельных частях ареала популяционная структура так же, как и у уральской лиственницы Сукачева, может быть определена как колониальная популяционная система, характеризующаяся относительной мономорфностью отдельных колоний, но в целом обладающая децентрализованным запасом генетической изменчивости [6].
Популяционной системе такого типа присущи некоторые собственные преимущества, основное из которых - существование в ней самой условий для быстрого микроэволюционного изменения [43]. Именно это имелось в виду, когда мы говорили о перспективах развития местной лиственницы Сукачева на Южном и Северном Урале за счет потенциала высокогорных популяций в случае благоприятного изменения условий существования. Еще в большей степени это следует отнести к высокополиморфным и уже сейчас имеющим тенденцию к распространению на восток приполярно-полярно-уральским популяциям.
Принципиально важным представляется сопоставление двух выявленных и подтверждаемых на основе генетического анализа фактов [26, 27, 34]: при низком уровне фенотипического и генетического разнообразия большинства уральских популяций лиственницы Сукачева (кроме крайне северных) степень ее межпопуляционной дифференциации оказывается не меньшей, чем у других хвойных видов. Следовательно, в эволюционном отношении факторы дифференциации на межпопуляционном уровне интенсифицированы здесь относительно в большей степени, чем микроэволюционные процессы в пределах популяций. Снижение фенотипического разнообразия внутри популяций в определенной степени компенсируется увеличением межпопуляционной изменчивости вида за счет «подключения» разнообразных эволюционных факторов.
Таким образом, обнаружено соответствие между популяционно-генетической структурой и микроэволюцией вида, с одной стороны, и историей расселения и современной структурой ареала лиственницы Сукачева в регионе -с другой. Генетический потенциал вида в обширной части уральского ареала рассредоточен по нескольким более или менее крупным локальным популяциям и их группам, различающиеся по фенотипическому и генетическому разнообразию и направлениям эволюционного развития.
Библиографический список
- Архипов С.А. Четвертичный период в Западной Сибири. Новосибирск: Наука, 1971. 331 с.
- Ваганов Е.А., Шиятов С.Г., Мазепа B.C. Дендроклиматические исследования в УралоСибирской Субарктике. Новосибирск: Наука, 1996.246 с.
- Горчаковский П.Л. История развития растительности Урала. Свердловск: Сверял, книж. изд-во, 1953. 143 с.
- Горчаковский П.Л. Флора и растительность высокогорий Урала: Тр. Ин-та биол. УФАН СССР. Вып. 48. Свердловск, 1966. 270 с.
- Горчаковский П.Л. Основные проблемы исторической фитогеографии Урала: Тр. Ин-та экол. раст. жив. УФАН СССР. Вып. 66. Свердловск, 1969. 286 с.
- Грант В. Эволюционный процесс. М.: Мир, 1991.488 с.
- Гричук В.П. История флоры и растительности Русской равнины в плейстоцене. М.: Наука, 1989. 183 с.
- Дылис Н.В. Сибирская лиственница. Материалы к систематике, географии и истории: Моск. об-во испыт. природы. Новая сер. Отдел ботан. Вып. 2. М.: Изд-во МОИП, 1947. 139 с.
- Животовский Л.А. Показатели популяционной изменчивости по полиморфным признакам // Фенетика популяций. М.: Наука, 1982. С. 38-44.
- Животовский Л.А. Популяционная биометрия. М.: Наука, 1991. 271 с.
- Игошина К.Н. Лиственница на Урале // Материалы по истории флоры и растительности СССР. Вып. 4. М.; Л.: Изд-во АН СССР, 1963. С. 462-492.
- Ильинский А.П. Методы и задачи изучения географического распространения древесных пород в СССР // Тр. по прикладной ботанике, генетике и селекции. Вып. 21. № 3. Л., 1929. С. 3-43.
- Кожевников Ю.П., Украинцева В.В. Некоторые особенности растительного покрова Евразии в древнем голоцене // Украин. бот. журн. 1992. Т. 77. №8. С. 1-9.
- Крашенинников И.М. Основные пути развития растительности Южного Урала в связи с палеографией Северной Евразии в плейстоцене и голоцене // Советская ботаника. 1939. № 6-7. С. 67-99.
- Кузнецов Н.И. Юго-западная, южная и юго-восточная граница распространения лиственницы (Larixsibirica Ledeb.) в пределах Европейской России // Юбилейный сборник, посвященный И.П.Бородину. Л., 1927. С. 231-256.
- Кулагин Ю.З. Лесообразующие виды, техногенез и прогнозирование. М.: Наука, 1980. 114 с.
- Куприянов Н.В. Лиственница Сукачева на юго-западной границе естественного распространения (Горьковское Заволжье) // Лиственница: Тр. Сиб. технол. ин-та. Т. 3. Красноярск, 1968. С. 44-51.
- Лузганов А.Г., Абаимов А.П. Роль речных бассейнов в расселении и эволюции лиственниц, кедра сибирского и других древесных пород // Лиственница (Межвуз. сб. науч. тр): Сиб. технол. ин-т. Т. 8. Красноярск, 1977. С. 31-38.
- Матвеев А.В. Эколого-генетическая изменчивость лиственницы сибирской (Larixsibirica Ldb.) на северном пределе ареала: Автореф. дисс. ... канд. биол. наук: 03.00.16. Екатеринбург: ИЭРиЖ УрО РАН, 1995. 16 с.
- Миллер Р., Кан Дж. Статистический анализ в геологических науках. М.: Мир, 1965. 482 с.
- Панова Н.К. История горных лесов центральной части Южного Урала в голоцене // Лесоведение. 1982. № 1. С. 26-34.
- Попов А.И. Четвертичный период в Западной Сибири // Ледниковый период на территории Европейской части СССР и Сибири. М.: Изд-во МГУ, 1959. С. 360-383.
- Попов Г.В. Основные этапы формирования растительного покрова Башкирского Предуралья в свете анализа реликтовых и эндемичных элементов флоры // Дикорастущие и интродуцируемые полезные растения в Башкирии. Вып. 3. Уфа, 1971. С. 254273.
- Попов Г.В. Леса Башкирии (их прошлое, настоящее и будущее). Уфа: Башкир, книж. издво, 1980. 144 с.
- Пугач Е.А. Изменчивость морфологических признаков у лиственницы Сукачева на Среднем Урале // Лиственница: Тр. Сиб. технол. ин-та. Сб. 39. Т. 2. Красноярск, 1964. С. 96-106..
- Путенихин В.П. Лиственница Сукачева на Южном Урале (изменчивость, популяционная структура и сохранение генофонда). Уфа: УНЦ РАН, 1993, 195 с.
- Путенихин В.П. Популяционная структура и сохранение генофонда хвойных видов на Урале: Автореф. дис. ... докт. биол. наук: 06.03.01. Красноярск: Ин-т леса им. В.Н. Сукачева СО РАН, 2000. 48 с.
- Райт Дж. В. Введение в лесную генетику. М.: Лесная промышленность, 1978. 470 с.
- Сукачев В.Н. К истории развития лиственниц // Лесное дело. М.; Л.: Новая деревня, 1924. С. 12-44.
- Сукачев В.Н., Поплавская Г.И. Очерк истории озер и растительности Среднего Урала в течение голоцена по данным изучения сапропелевых отложений // Бюлл. Комис. по изуч. четвертич. периода. № 8. М.; Л.: Изд-во АН СССР, 1946. С. 5-37.
- Тараканов СВ. Генетическая гетерогенность ценопопуляций ели Шренка в экологическом оптимуме и пессимуме // Экологические исследования биоты экосистем Северной Киргизии. Фрунзе, 1988. С. 23-30.
- Тимофеев-Ресовский Н.В., Воронцов Н.Н., Яблоков А.В. Краткий очерк теории эволюции. М.: Наука, 1969. 408 с.
- Чигуряева А.А., Хвалина Н.Я. Материалы к флоре и растительности Башкирского Предуралья в голоцене // Науч. докл. высш. школы (биол. науки). 1961. № 1. С. 131-138.
- Шигапов З.Х., Путенихин В.П., Шигапова А.И., Уразбахтина К.А. Генетическая структура уральских популяций лиственницы Сукачева // Генетика. 1998. Т. 34. № ]. С. 65-74.
- Юдин Ю.П. Распространение лиственницы между Печорой и Уралом // Известия Коми фил. Всесоюз. географ, об-ва. Вып. 3. Сыктывкар, 1955. С. 28-33.
- Dobzhansky T. Genetics of evolutionary processes. N.Y.: Columbia Univ. Press, 1970. 505 pp.
- Fins L., Seeb L.W. Genetic variation in allozymes of western larch // Can. J. Forest Res. 1986. V. 16. N5. P. 1013-1018.
- Kullman L. Palaeoecological, biogeographical and palaeoclimatological implications of early Holocene immigration of Larix sibirica Ledeb. into the Scandes Mountains, Sweden // Global Ecology and Biogeography Letters. 1998. V.7.N3.P. 181-188.
- Park Y.S., Fowler D.P. Effects of inbreeding and genetic variances in a natural population of tamarack (Larix laricina (Do Roi) K. Koch in eastern Canada // Silvae Genet. 1982. Bd. 31. H. l.S. 21-24.
- Parker W.H., Dickinson T.A. Range-wide morphological and anatomical variation in Larix laricina II Can. J. Bot. 1990. V. 68. N 4. P. 832-840.
- Rehfeldt G.E. Genecology of Larix laricina (Du Roi) K. Koch in Wisconsin // Silvae Genet. 1970. Bd 19. H. l.S. 9-16.
- Rehfeldt G.E. Differentiation of Larix occidentalis populations from the Northern Rocky Mountains// Silvae Genet. 1982. Bd 31. H. l.S. 1319.
- Wright S. Evolution in Mendelian populations//Genetics. 1931. V. 16. P. 97-159.
|


