|
Влияние стволовой гнили на энтоморезистентность березы повислой (Betula pendula) в системе: береза-непарный шелкопряд
Колтунов Е.В., Клобуков Г.И.
Ботанический сад Уральского отделения РАН
Екатеринбург
Установлено, что при значительном диаметре поражения березы повислой стволовой гнилью, превышающем 30% от диаметра ствола, уровень дефолиации крон этих деревьев гусеницами непарного шелкопряда заметно возрастает. В составе древостоев порослевого происхождения доминировали деревья с наибольшей степенью поражения ствола, тогда как в составе популяции березы семенного происхождения, наоборот, деревьев с сильным поражением ствола гнилью было минимальное количество, а доминировали деревья либо здоровые, либо очень слабо пораженные (0-15%). Это свидетельствует о том, что березняки порослевого происхождения имеют пониженный иммунитет к грибам, вызывающим стволовые гнили. Учитывая чрезвычайно высокий уровень распространения стволовой гнили в березовых лесах Зауралья (70-90%) вследствие ослабленности лесов антропогенными факторами, а, также, доминирование древостоев порослевого происхождения, этот фактор, наряду с возможными климатическими сдвигами, может служить заметным фактором роста площади очагов массового размножения насекомых-филлофагов и интенсивности вспышек.
Ключевые слова: стволовая гниль, березы повислая, непарный шелкопряд, энтоморезистентность
Crones defoliation level of birch drooping by Lymantria dispar L. considerably increases, that at considerable diameter of defeat of trees by rotting of stem, exceeding 30 % from stem diameter, it is established. As a part of forest stands of second growth origins dominated trees with the greatest degree of stem affection whereas as a part of population of a birch of a seed origin, on the contrary, trees with strong of stem affection by rotting of stem there was a minimum quantity, and trees either healthy, or very poorly affection (0-15 %) dominated. It testifies that birch forests of second growth origins have the lowered immunity to the mushrooms causing rotting of stem. Considering extremely high level of rotting of stem distribution in birch forests of the Urals (70-90 %) in consideration of attenuated of stands by anthropogenous factors, and, also, domination of second growth origins forest stands, this factor, along with possible climatic shifts, can serve as the appreciable factor of growth of the area of insects-phytophages outbreaks and intensity of outbreaks.
Key words: Rotting of stem, birch drooping, entomoresistance, Lymantria dispar L.
Уровень и масштабы антропогенного воздействия на лесные биоценозы постоянно возрастают. При этом часто высокие рекреационные нагрузки сочетаются с техногенным загрязнением лесов. Это сопровождается снижением их устойчивости к насекомым-вредителям и инфекционным болезням. В ослабленных лесах особенно широкое распространение получили очаги массового размножения насекомых-филлофагов и корневые и стволовые гнили (Колтунов, 1993, Колтунов и др., 1998, 2007). В этой ситуации особенно актуален вопрос о влиянии корневых и стволовых гнилей на уровень энтоморезистентности древостоев. До настоящего времени этот вопрос остается, практически, неизученным. Ранее нами обнаружено, что бактериальная водянка значительно снижала энтомотолерантность березы повислой в южном Зауралье (Колтунов, 1993, Колтунов и др., 1998). Эта проблема имеет важное теоретическое и практическое значение, так как снижение устойчивости пораженных гнилями древостоев может служить одним из факторов роста площади очагов массового размножения насекомых-филлофагов.
Материалы и методы. Исследования проводили в очагах массового размножения зауральской географической популяции непарного шелкопряда в северной части ареала, в березовых лесах Каменск-Уральского района Свердловской области. В этих лесах наблюдается периодическое возникновение вспышек массового размножения этого фитофага (Колтунов, 1993; Колтунов и др., 1998), а, также – насекомых-филлофагов летне-осеннего комплекса. В 2006 г. возникла очередная вспышка массового размножения этого фитофага. Вследствие неблагоприятных климатических условий (низкие зимние температуры декабря, вызвавшие гибель кладок) и недостаточно благоприятных условий весенне-летнего периода отрождаемость яиц и выживаемость популяции была незначительной. Средний уровень дефолиации крон составлял 15-30%, у отдельных деревьев до 50-80%. Непосредственно перед определением степени дефолиации крон березы на пробных площадях проводили отбор кернов из ствола приростным буравом и определение наличия, или отсутствия стволовой гнили. Кроме того идентифицировали, также, ширину зоны поражения ствола гнилью (по отношению ширины инфицированной древесины керна к общей его длине). Одновременно этот параметр служил, также, показателем стадии развития болезни. Затем деревья одного диаметра, растущие в непосредственной близости, в одинаковых лесорастительных условиях, с одинаковой заселенностью кладками разделялись на контрольные (здоровые) и опытные (с признаками стволовой гнили, с разной шириной инфицированной зоны). Для количественной оценки степени дефолиации крон березы использовали бинокль. Проводились отдельные измерения разных (по высоте) частей кроны (верхняя, средняя, нижняя). В каждом варианте было по 20 деревьев. Количественный анализ степени дефолиации крон проводился нами в разные сроки (с июня – до завершения периода активного питания гусениц и окукливания). Статистическую обработку полученных данных проводили с использованием Т-критерия Cтьюдента при Р? 0,05.
Как показали результаты, у березовых древостоев в районе очагов выявлен очень высокий процент пораженности стволовой гнилью. Так в разной степени в Покровском лесничестве поражены 70-80% деревьев, а в п. Кисловское это значение достигло 90-100%. В первом варианте средний диаметр пораженной зоны (относительно общего диаметра ствола) составлял 18,3%. При этом значения варьировали от 0% до 63%, во втором это значение было, в среднем, выше: 20,2%, при варьировании от 1,4% до 94%.
Для изучения влияния стволовой гнили на степень дефолиации крон березы была проведена сравнительная оценка степени дефолиации опытных (пораженных стволовой гнилью), в зависимости от ширины зоны инфицирования, и контрольных деревьев (здоровых) на пробных площадях. Результаты исследований убедительно показали, что средние значения дефолиации крон (общая дефолиация) в вариантах с максимальной степенью пораженности стволовой гнилью (>30%) и остальными достоверно различались (рис. 1). Средние значения дефолиации различались, в среднем, в два раза. Относительно низкие параметры максимальной дефолиации обусловлены как неблагоприятными климатическими условиями, так и активацией фактора индуцированной резистентности у березняков, вследствие дефолиации в предыдущем году не менее чем на 30%.
Кроме того, были изучены, также, особенности дефолиации верхней части крон березы у пораженных стволовой гнилью деревьев (в зависимости от степени развития болезни (ширины инфицированной зоны древесины ствола)) и здоровых. Как показали результаты, наиболее значительные различия в степени дефолиации верхней части крон имели место между наиболее пораженными стволовой гнилью деревьями (>30%, в среднем – 68%), и здоровыми и слабо пораженными (рис. 2). Различия статистически достоверны. Между контрольными вариантами, слабо - и средне пораженными статистические различия недостоверны.
Учитывая полученные нами результаты об исключительно высоком уровне распространения стволовой гнили в березовых лесах Каменск-Уральского района в очагах было вполне обоснованным связать этот процесс с большим процентом березы порослевого происхождения, как показано нами ранее (Колтунов, 1993). Исходя из этого, нами было изучено, также, влияние происхождения березы (порослевое, семенное) на степень пораженности ствола гнилью (процентное соотношение ширины зоны инфицированной древесины керна к общей его длине). По существу, этот параметр отражает стадию развития заболевания, интенсивность развития болезни, которые обусловлены, главным образом, уровнем иммунитета дерева.
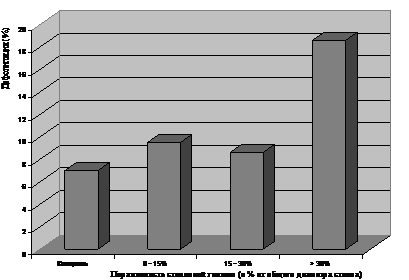
Рисунок 1 - Влияние степени пораженности березы стволовой гнилью на уровень дефолиации кроны непарным шелкопрядом
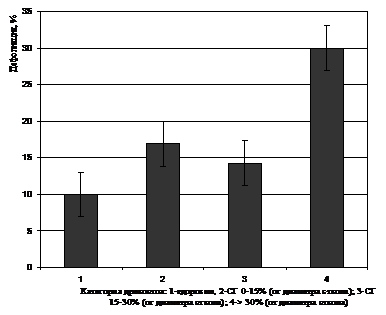
Рисунок 2 – Степень дефолиации верхней части кроны березы повислой в зависимости от уровня пораженности стволовой гнилью
Результаты исследований убедительно показали, что в составе древостоев порослевого происхождения доминировали деревья с наибольшей степенью поражения ствола, тогда как в составе популяции березы семенного происхождения, наоборот, деревьев с сильным поражением ствола гнилью было минимальное количество, а доминировали деревья либо здоровые, либо очень слабо пораженные (0-15%) (рис. 3). Из этого же графика отчетливо видно, что в составе семенных деревьев березы здоровых экземпляров было почти в два раза больше, чем среди порослевых, хотя средний уровень заболевания среди каждой из подгрупп был высоким (1-66,6%; 2-81,82%). Это свидетельствует о том, что березняки порослевого происхождения, очевидно, имеют пониженный иммунитет к грибам, вызывающим стволовые гнили. Поэтому они в большей степени поражаются этим патогеном, при этом, очевидно, возрастает и интенсивность развития болезни, которая отражается на общем диаметре поражения стола гнилями.
Таким образом, результаты исследований влияния стволовой гнили березы повислой на степень дефолиации крон убедительно показали, что при значительном диаметре поражения ствола гнилью, превышающем 30% от диаметра ствола, уровень дефолиации крон этих деревьев гусеницами непарного шелкопряда заметно возрастает. Мы предполагаем, что это обусловлено снижением уровня энтоморезистентности березняков, значительно пораженных стволовой гнилью. Это может происходить как за счет благоприятных изменений биохимического состава кормового субстрата (листьев), так и за счет снижения активности синтеза различных антифидантных соединений. Как показали результаты исследований, у березы повислой порослевого происхождения выявлено более значительное снижение уровня иммунитета к заболеванию стволовой гнилью, по сравнению с древостоем семенного происхождения, вследствие чего происходит возрастание как интенсивности развития болезни, так и распространенности ее среди этой группы древостоев. Учитывая чрезвычайно высокий уровень распространения стволовой гнили в березовых лесах Зауралья вследствие ослабленности лесов антропогенными факторами, а, также, доминирования древостоев порослевого происхождения, этот фактор, наряду с возможными климатическими сдвигами, может служить заметным фактором роста площади очагов массового размножения насекомых-филлофагов и интенсивности вспышек. Учитывая это, совершенно очевидно вытекает целесообразность постепенной реконструкции березовых лесов и замены порослевых березняков семенными.
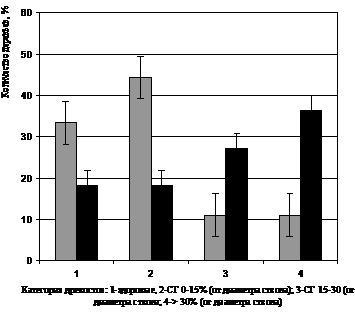
Рисунок 3 – Влияние происхождения деревьев (1-семенное, 2-порослевое) на пораженность стволовой гнилью и степень развития болезни (от общего диаметра ствола)
БИБЛИОГРАФИЧЕСКИЙ СПИСОК
Колтунов, Е.В. Корневая и стволовая гнили сосны обыкновенной (Pinus sylvestris) в городских лесопарках г. Екатеринбурга / Е.В. Колтунов, C.В. Залесов, Лаишевцев Р.Н. // Леса России и хозяйство в них: сб. ст. – Екатеринбург, Изд-во УГЛТУ и БС УрО РАН, вып. 1, 2007, С. 238-246.
Колтунов, Е.В. Насекомые-фитофаги лесных биогеоценозов в условиях антропогенного воздействия / Е.В. Колтунов – Екатеринбург: Наука, 1993. -137 с.
Колтунов, Е.В.. Экология непарного шелкопряда в условиях антропогенного воздействия. Екатеринбург / Е.В. Колтунов, В.И. Пономарев, С.И. Федоренко – Екатеринбург, Изд-во УрО РАН, 1998. - 214 с.
|


